接種時期選定の参考 (新)

ここでは「3-1-2-2.抗体(1)」で紹介した「B細胞が1人前の抗体産生細胞になるまでに受ける試練」の内容をもう少し詳しく見ていきます。(仮説です)
| 試練の場所 | チェックの内容 |
| 骨髄 | 作ったものがちゃんと抗体の形をしていること |
| 自己抗原とは結合しないこと | |
| リンパ節 | 病原体の抗原としっかり結合すること |
抗体の設計図に相当するもの(アミノ酸の配列順序を決めるもの)は 遺伝子 です。
細胞の中に 核 と呼ばれる部分があり、細胞核の中に DNA があります。
そのDNAのあちらこちらに、細胞の作るタンパクのアミノ酸配列の情報(これが遺伝子:タンパクの設計図)が保持されています。
細胞核のDNAは細胞分裂の時はいくつかのかたまり(染色体)を形成します。
では、「3-1-2-2.抗体(1)」で見た抗体の姿を思い出しましょう。
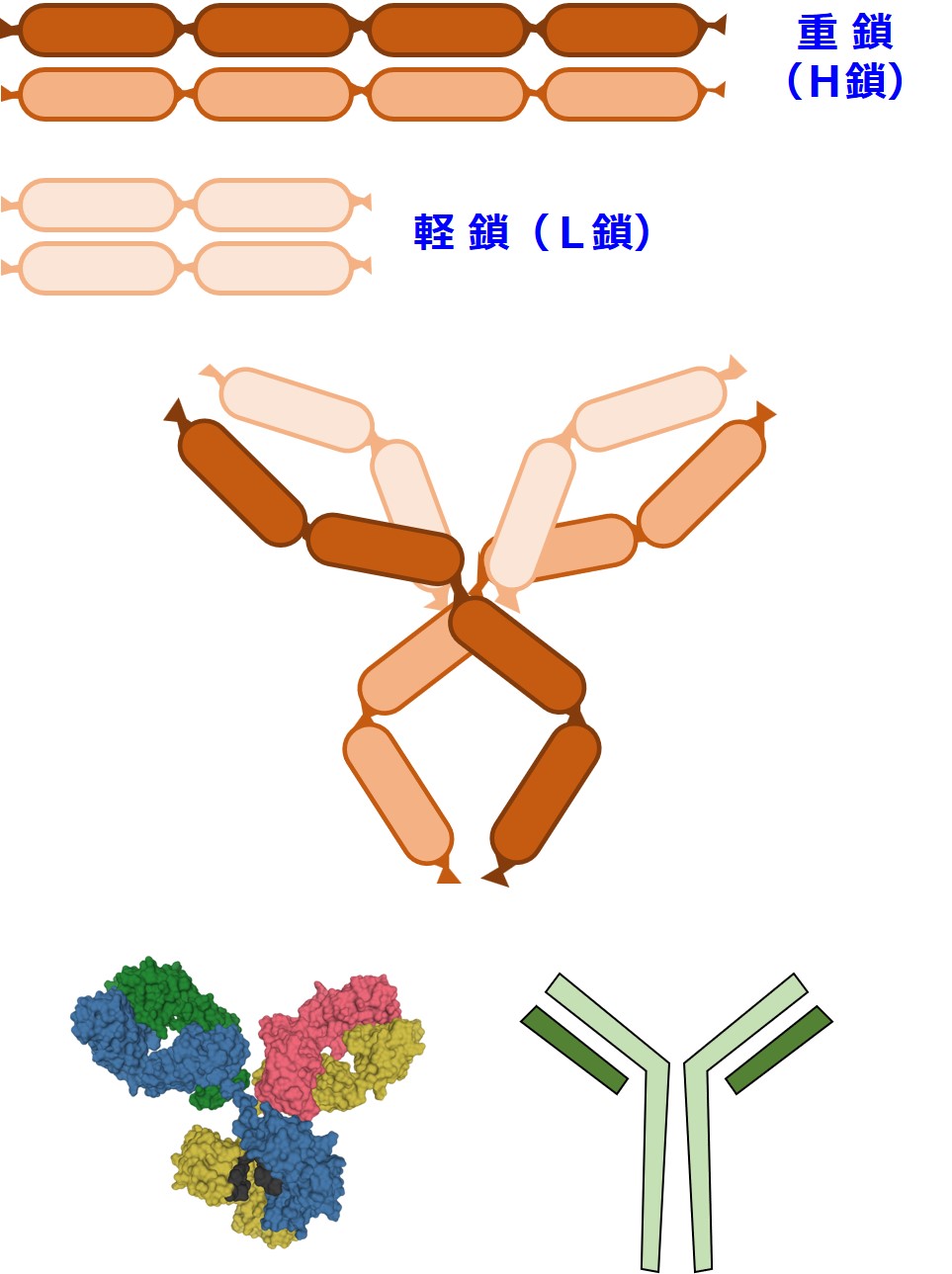
抗体の重鎖(H鎖)の設計図と軽鎖(L鎖)の設計図は、別々の染色体にあります。つまり、ヒトのDNA上の遠く離れたところにあるということです。
したがって、重鎖(H鎖)と軽鎖(L鎖)は生産時に設計図の共用はしていないと考えられています。
H鎖とL鎖は別々に作られて合体するようです。
ということで、重鎖(H鎖)の生産と軽鎖(L鎖)の生産を別々に見ていかないといけません。
ここでは、先に重鎖(H鎖)作りと軽鎖(L鎖)作りのことを少しだけ紹介し、それから抗体作りの全体像を示します。
全体の流れを示した後で、重鎖(H鎖)作りと軽鎖(L鎖)作りを並行で細かく説明します。
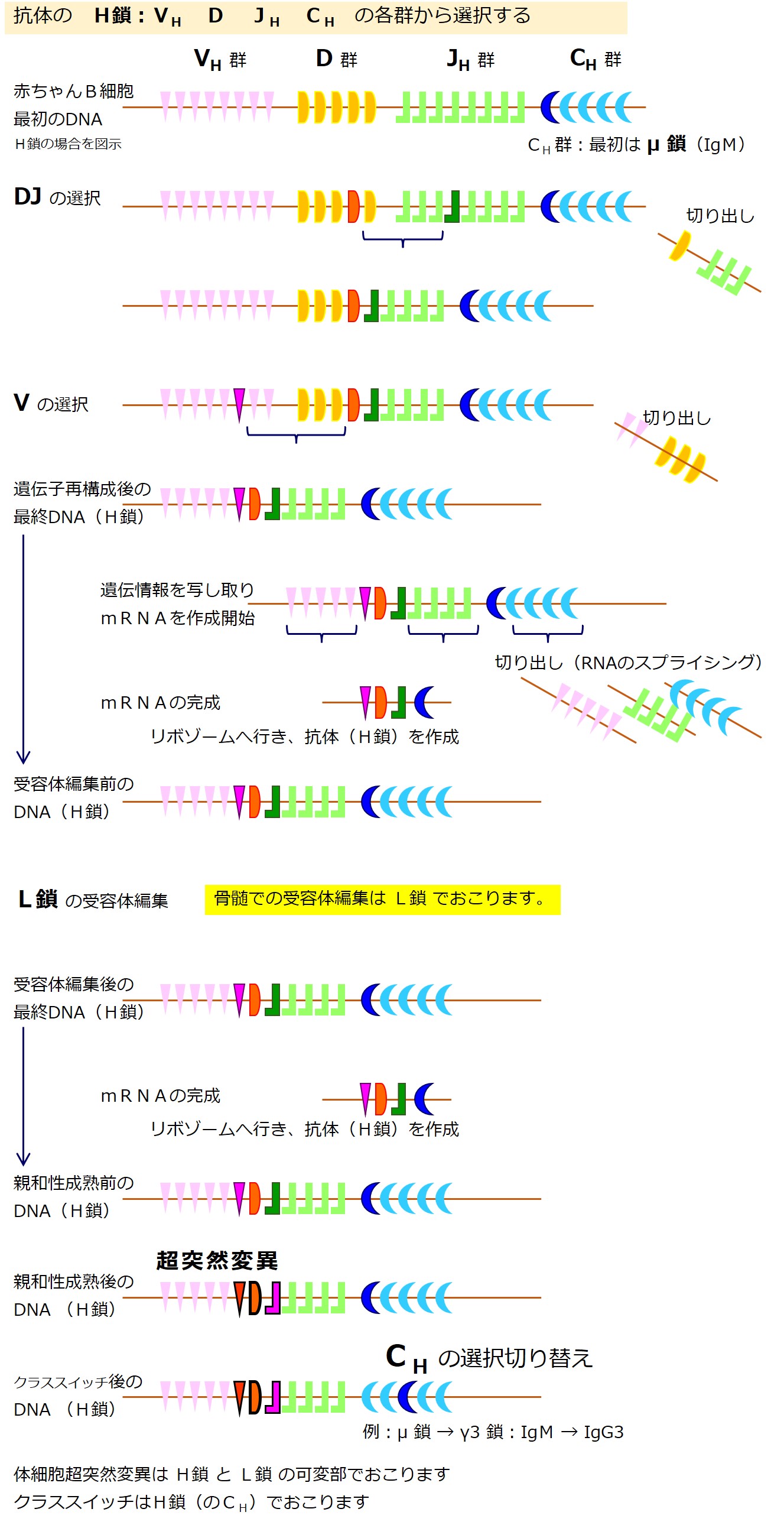
では、次の図を使って抗体の重鎖(H鎖)の構成について説明します。
知らない初めての名前がいっぱいあるかもしれませんが、もちろん、覚えなければならない名前は何ひとつありません。
抗体がどのような流れで作られるのかが理解できればそれでよいのです。流れが分かった後で、名前などは全部忘れてかまいません。
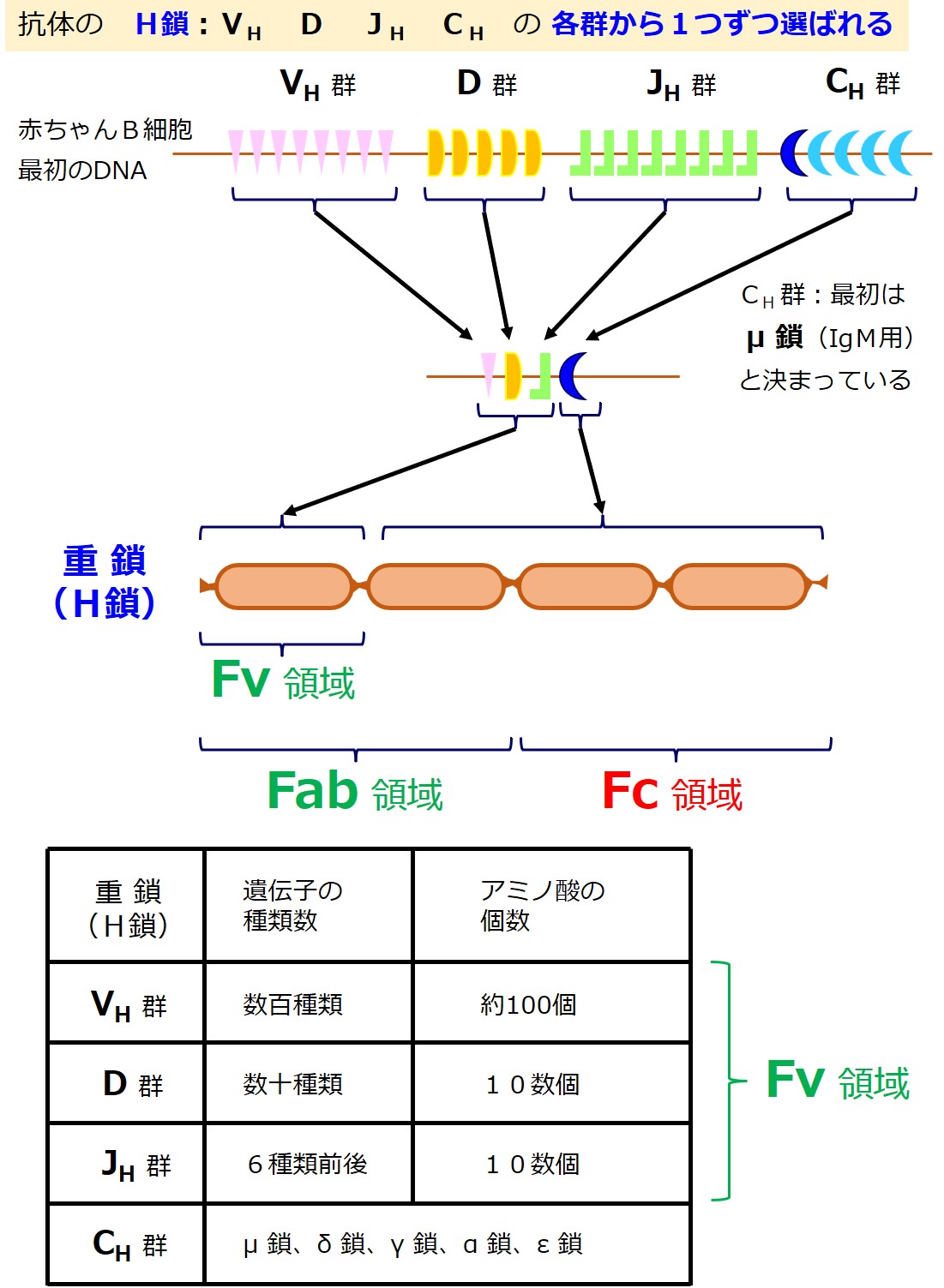
上図を説明します(図はあくまで便宜的イメージであり、遺伝子の数などは不正確です)。
重鎖(H鎖)は、下図のように4つのドメイン(お肉のかたまり)を形成しています。
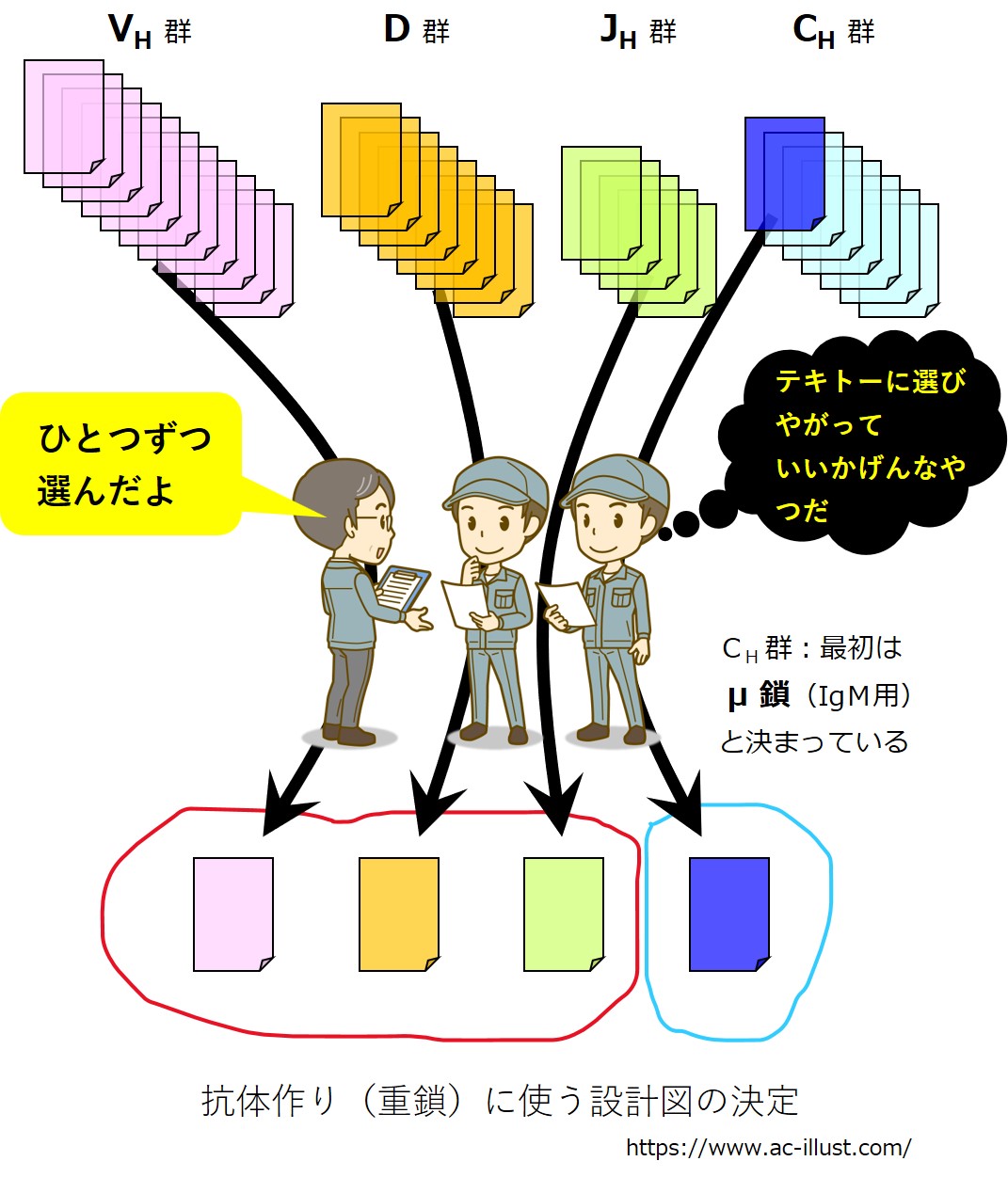
そのうち、抗原(エピトープ)と結合するドメイン(Fv 領域)のアミノ酸配列を決める遺伝子は、V、D、Jの3種類から構成されています。
それは何百種類とある VH 群の遺伝子の中からひとつ選び出され、同様に数十種類あるD群の遺伝子の中からひとつ選び出され、同様に6種類前後ある JH 群の遺伝子の中からひとつ選び出され、「V-D-J」の3つで構成されます。
抗原(エピトープ)と結合するドメイン(Fv 領域)以外の3つのドメインのアミノ酸配列を決める遺伝子は、Cの1種類であり、CH 群の遺伝子の中からひとつ選び出されます。
最初に作られる抗体の重鎖(H鎖)では、Cは μ 鎖(ミュー鎖)と決まっています(最初に作られる抗体は IgM になります)。
重鎖(H鎖)全体では、「V-D-J-C」とつながった遺伝子が作られます。
この遺伝子作成は、たくさんの遺伝子を持つDNAを切ったり繋げたりして行われるので「遺伝子再構成(遺伝子編集)」と呼ばれます。
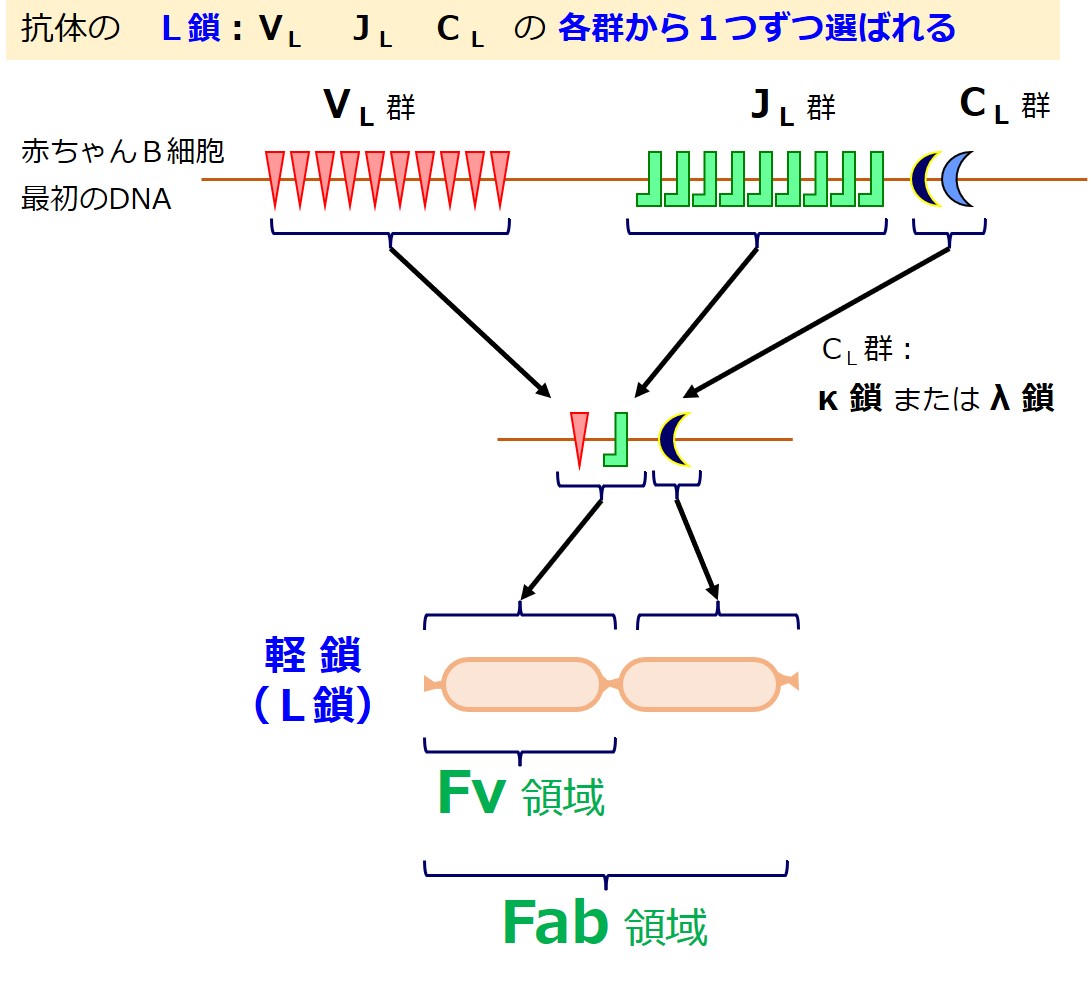
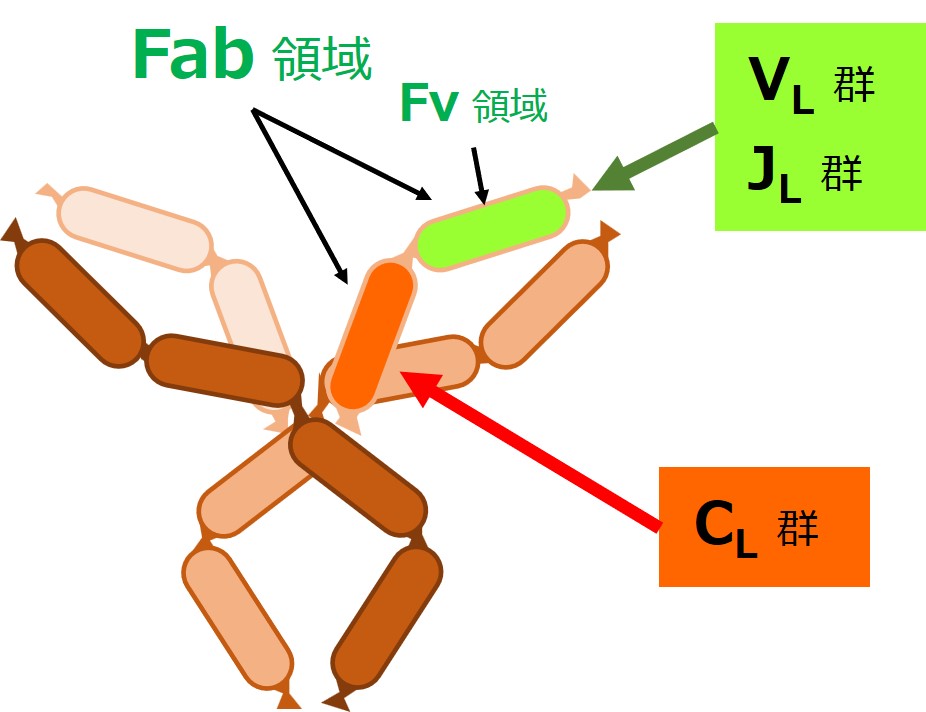
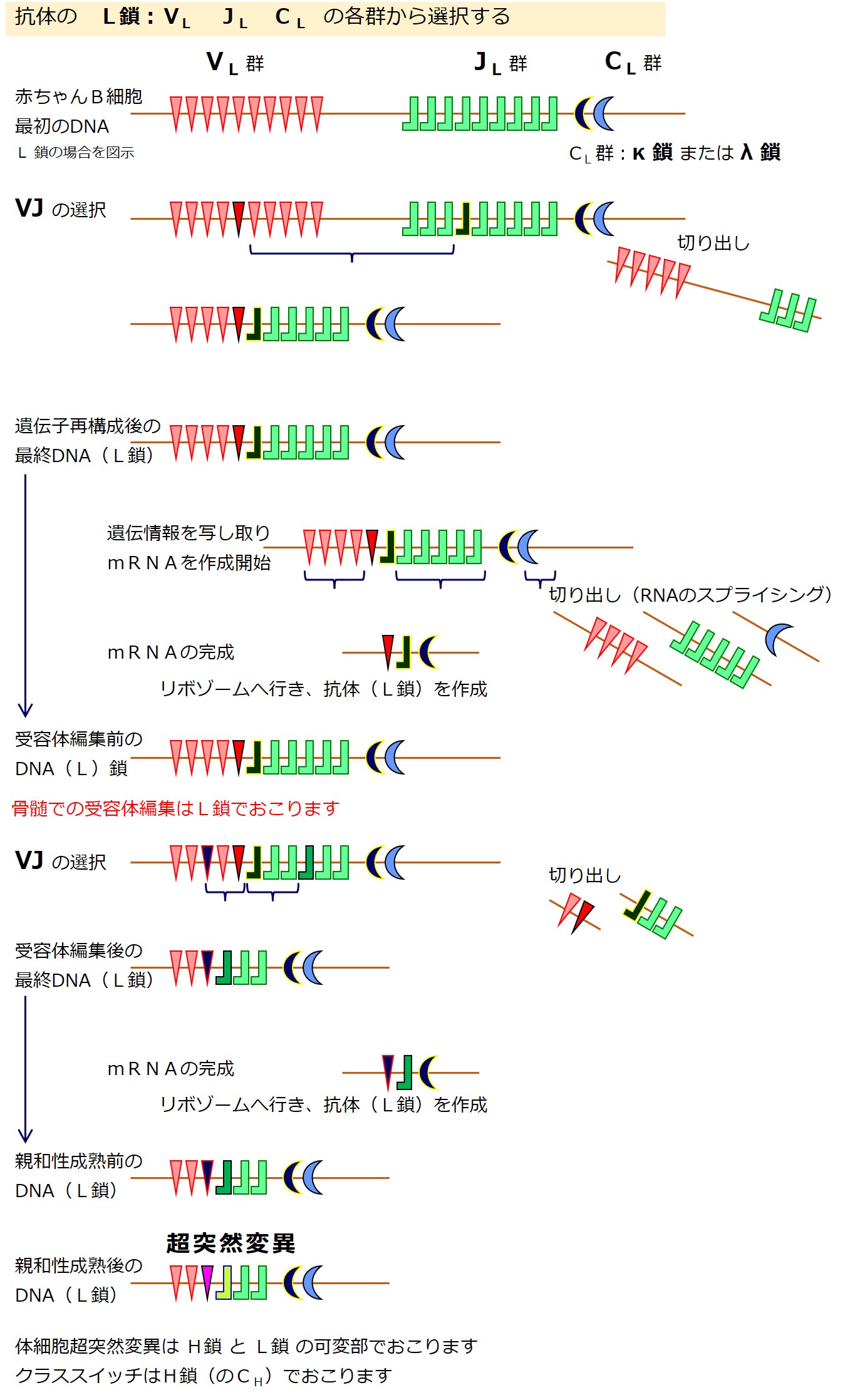
抗体の軽鎖(L鎖)の場合は、次図のようにD群遺伝子はありません。

ではここで、抗体作りの全体図を示します。


以上の流れを表にすると下表のようになります。
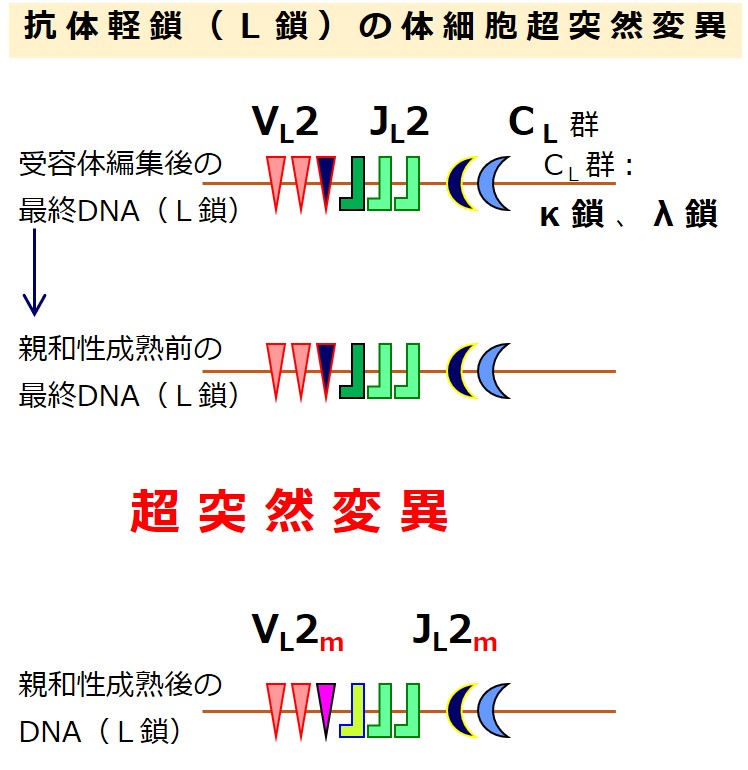
表では次のように表記しています:{VL群}の中から選ばれた最初の設計図を「VL1」、(骨髄での受容体編集で)次に選ばれた設計図を「VL2」、(リンパ節での親和性成熟つまり体細胞高頻度突然変異で)書き換えられた設計図を「VL2m」、もう一度書き換えられた設計図を「VL2m2」、さらに書き換えられたときは「VL2m3」。
この表などは、テキトーに眺めて流れをつかむだけでかまいません。
| ★ 骨髄において | |||
| 重鎖(H鎖) | 軽鎖(L鎖) | 検査内容 | |
| 変化 | 重鎖遺伝子再構成 | ||
| 構成元 | {VH群, D群, JH群, CH群} | ||
| 新構成 | VH1-D1-JH1-CH1 | ||
| 審査 | 仮軽鎖を使って抗体を作成し受検 | 抗体の体をなしているか? | |
| 選別 |
|
H鎖作りが合格してからL鎖作りが始まります | |
| 変化 | (H鎖に変化なし) | 軽鎖遺伝子再構成 | |
| 構成元 | {VL群, JL群, CL群} | ||
| 新構成 | VL1-JL1-CL1 | ||
| 審査 | 受検 | 自己抗原とは結合しないか? | |
| 選別 |
|
||
| 変化 | (H鎖に変化なし) | 受容体編集(軽鎖) | |
| 新構成 | VL2-JL2-CL1 | ||
| 審査 | 受検 | 自己抗原とは結合しないか? | |
| 選別 |
|
||
| 変化 | (H鎖に変化なし) | 受容体編集(軽鎖) | |
| 新構成 | VL3-JL3-CL1 | ||
| 選別 | 受検:合格・・・としておく | 自己抗原とは結合しないか? | |
| 合格したB細胞は骨髄を出て全身を循環 → 感染時はリンパ節に集まる | |||
| ★ リンパ節において | |||
| B細胞受容体に結合した病原体を食べて少し活性化する | |||
| 同じ病原体による活性化ヘルパーT細胞の刺激を受けてB細胞は大いに活性化し増殖する | |||
| 審査 | 受検 | 病原体抗原との結合 | |
| 選別 |
|
||
| 変化 | 1回目の親和性成熟(体細胞超突然変異) | ||
| 新構成 | VH1m-D1m-JH1m-CH1 | VL3m-JL3m-CL1 | |
| 変化 | 同じ頃に クラススイッチ もおこる(H鎖のCH) | ||
| 新構成 | VH1m-D1m-JH1m-CH2 | (L鎖に変化なし) | |
| 審査 | 受検 | 病原体抗原との結合 | |
| 選別 |
|
||
| 変化 | 2回目の親和性成熟(体細胞超突然変異) | ||
| 新構成 | VH1m2-D1m2-JH1m2-CH2 | VL3m2-JL3m2-CL1 | |
| 審査 | 受検 | 病原体抗原との結合 | |
| 選別 |
|
||
| 変化 | 3回目の親和性成熟(体細胞超突然変異) | ||
| 新構成 | VH1m3-D1m3-JH1m3-CH2 | VL3m3-JL3m3-CL1 | |
| 審査 | 受検 | 病原体抗原との結合 | |
| 感染症が治り、病原体の数が減るとこうした親和性成熟自体ができなくなり、抗体の改良は終了する。 | |||
遺伝子に加わる変化を順番に示すと次図になります。
各段階の少し詳しい説明はあとでおこないますので、まずは図をさっと眺めて下さい。
まず重鎖(H鎖)の場合:
次に軽鎖(L鎖)の場合:
・・・工事中・・・各段階の説明を加える・・・予定
ここまでの説明で、概略はつかめたと思われます。
では、一人前の立派な抗体産生が始まるまでを、各段階ごとに少し詳しく説明していきます。
骨髄で生まれた幼若なB細胞(赤ちゃんB細胞)は、喰って寝て、喰って寝てを繰り返しながら成熟し、そのまま血液中に出て行って働くわけではありません。
骨髄で生まれた幼若なB細胞の9割以上は骨髄で殺されてしまうようです。
もったいないですよね。どうしてそのようなことになるのでしょう。
抗体というのは、極めて強力な武器です。
爆薬、化学兵器、生物兵器、核兵器を弾頭に積載できる精密誘導ミサイルのようなものです。
たとえば、IgGに捕まった抗原は多種多様な攻撃を受け、破壊によって排除されます。
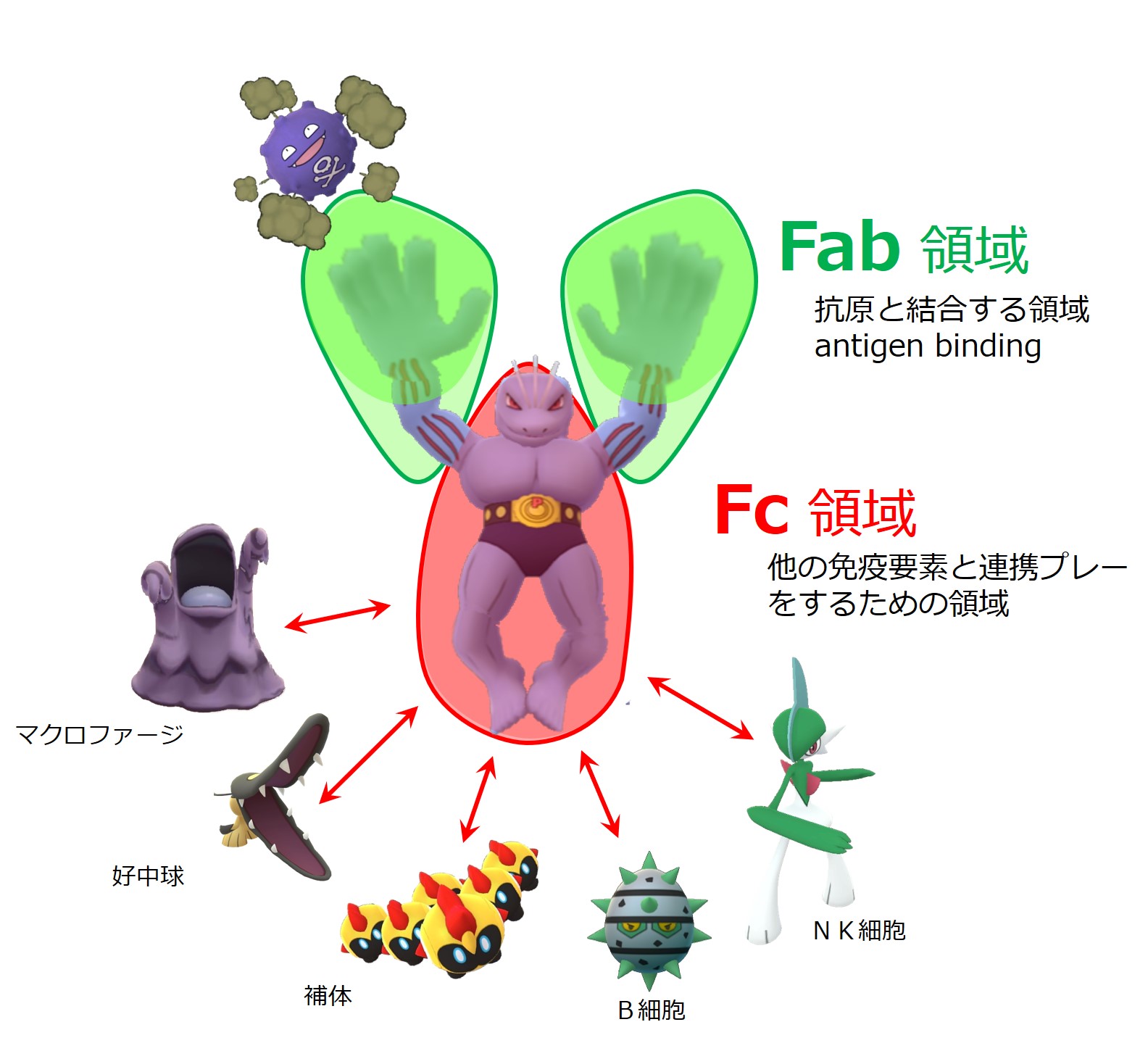
抗体にピタッと掴まえられると抗原は「ひぇ~!」とビビるわけです。
それだけに、ちゃんとした抗体を作れないとき、B細胞は殺されてしまうのです。
ちゃんとした抗体に求められる最も重要な性能は、「仲間を攻撃しない」という性能です。
その性能を確かに持っている抗体を産生できると確認されたB細胞だけが、骨髄を出て全身を巡回警備する任務に就きます。
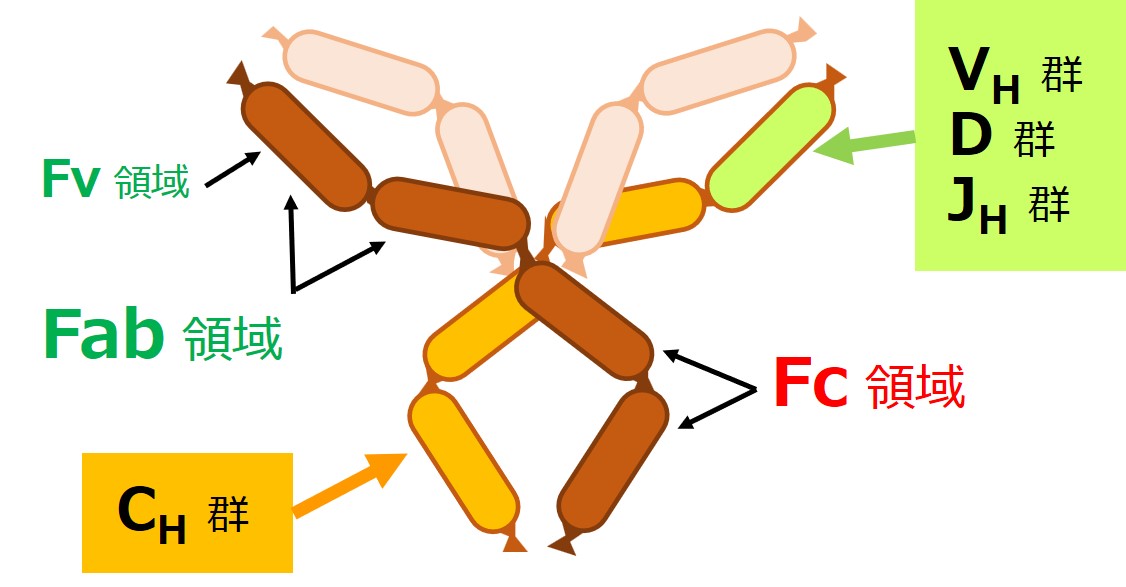
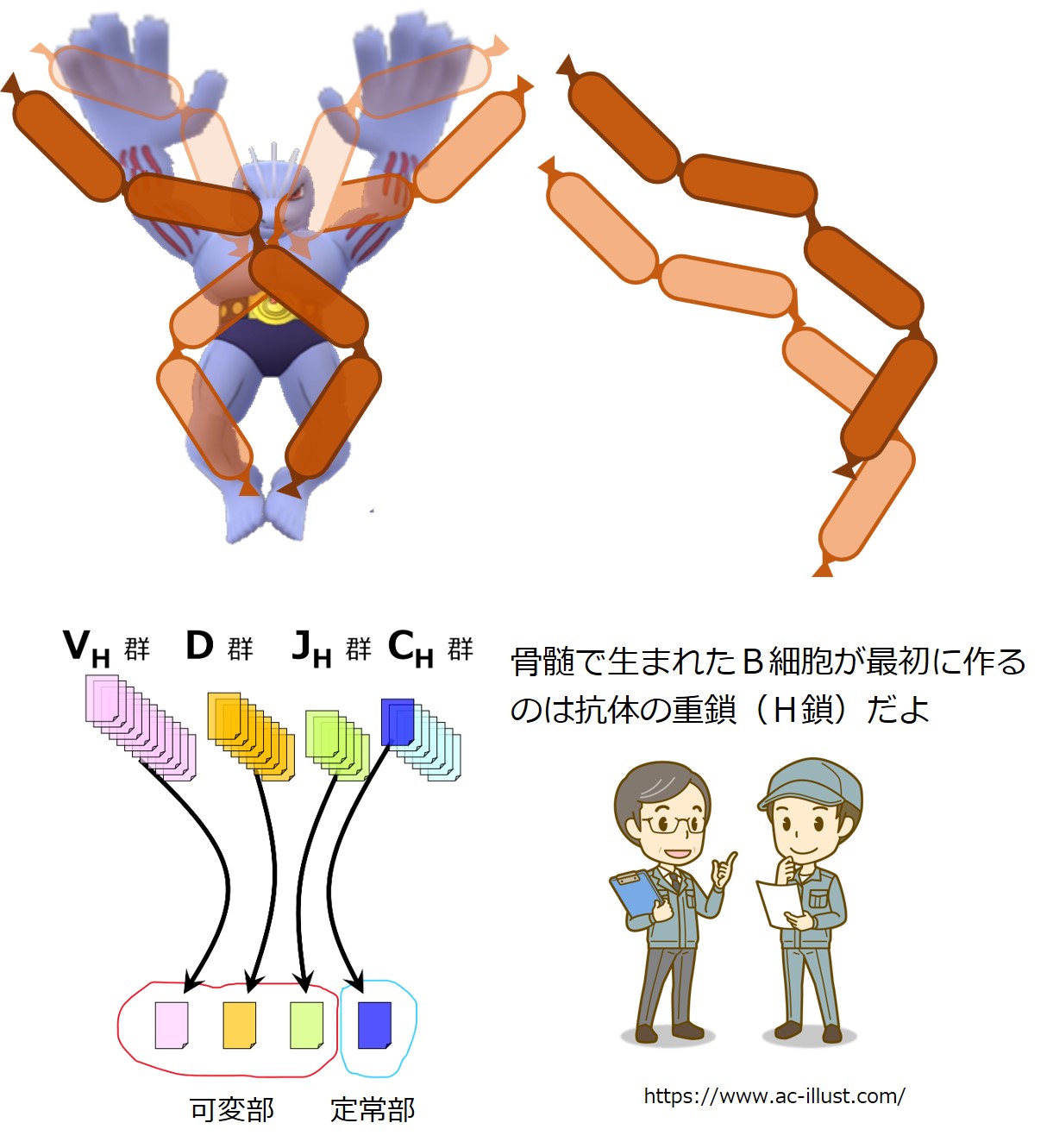
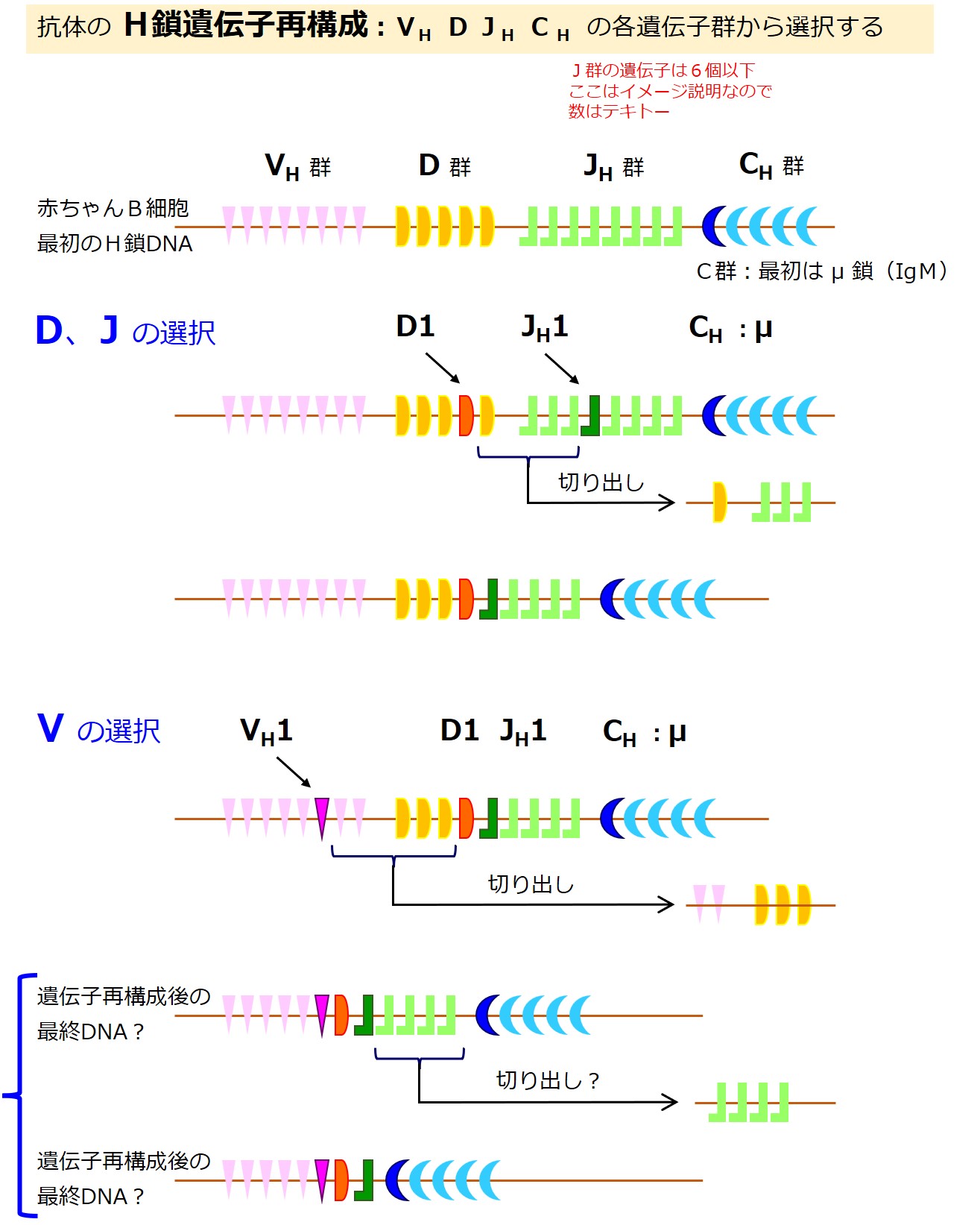
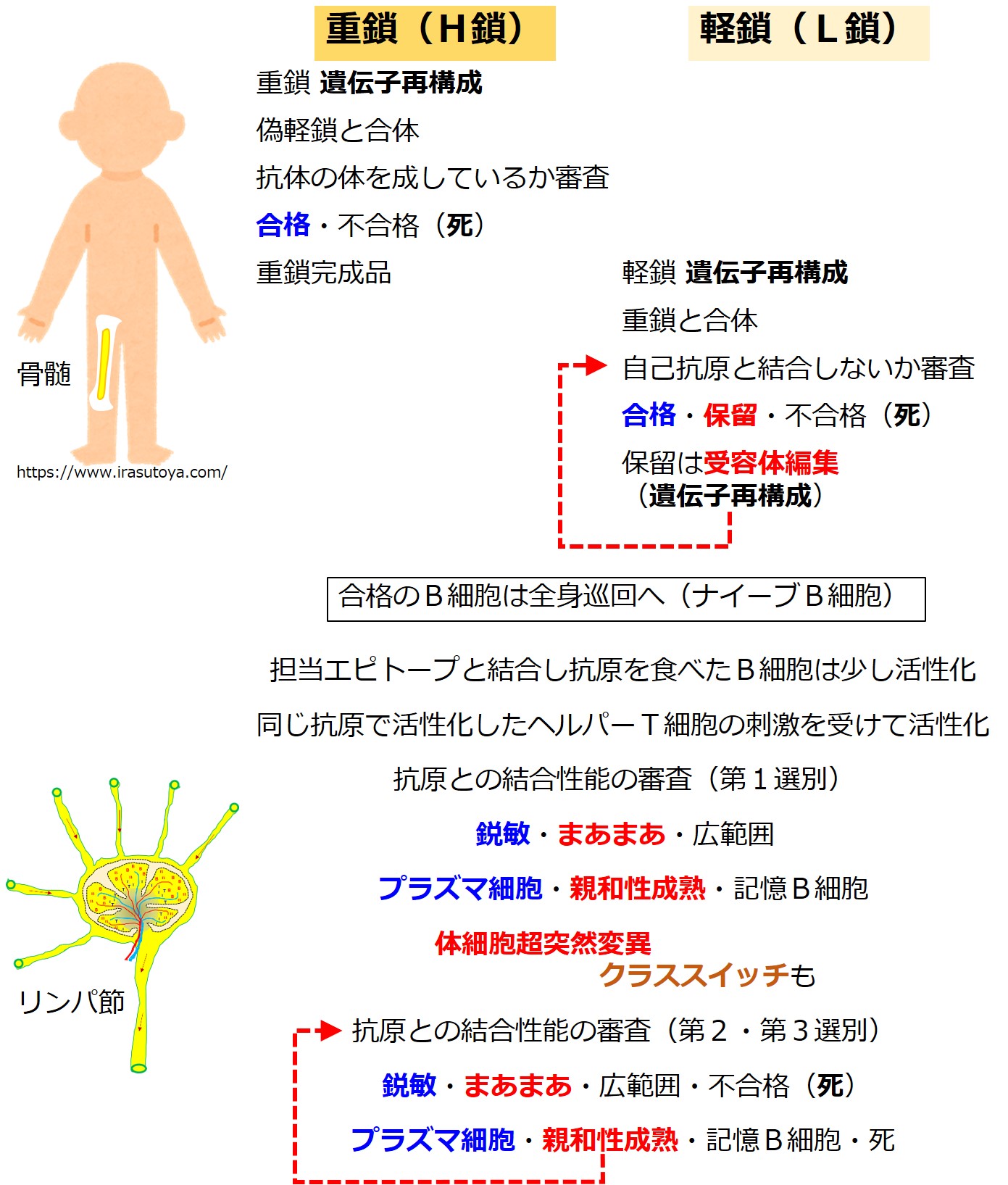
赤ちゃんB細胞が最初に作るのは抗体の重鎖(H鎖)です。
重鎖(H鎖)が抗体のどの部分か、再確認しましょう。
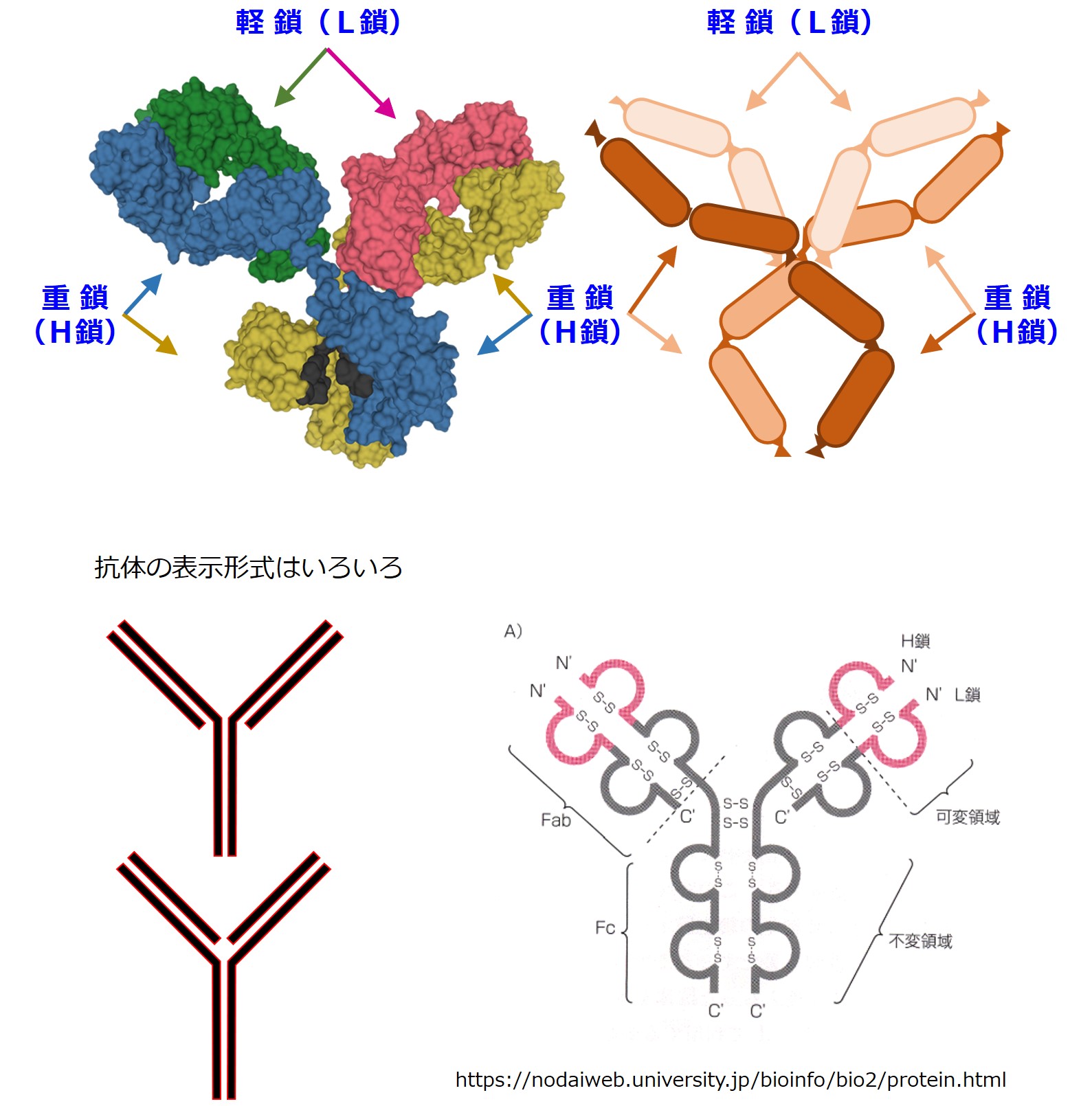
右下図は東京農業大学のウェブページ「タンパク質の立体構造」より。RASwin を使って IgG をいろんな方法で表現できます。
重鎖(H鎖)の遺伝子は、{V、D、J、C}の各遺伝子群に分かれています。
軽鎖(L鎖)にはD群が無いので、重鎖(H鎖)の遺伝子群は{VH-D-JH-CH}と表現されることがあります。
VH 群には、約100個のアミノ酸配列情報を持つ遺伝子が数百種類あります。
D 群には、約10数個のアミノ酸配列情報を持つ遺伝子が数十種類あります。
JH 群には、約10数個のアミノ酸配列情報を持つ遺伝子が6種類前後あります。
CH 群には、約400個~500個以上(4~5個のドメイン数)のアミノ酸配列情報を持つ遺伝子が 10 数種類あります(μ、δ、γ、α、ε、とそれらのサブクラス)。
幼若なB細胞は、自分が産生する抗体を1種類決めなければなりません。
自分はこれを産生すると決めた抗体の良し悪しで、B細胞が生きるか死ぬかが決まります。
最初に重鎖(H鎖)に用いる遺伝子を決めます。
重鎖(H鎖)に用いる遺伝子の選択チャンスは一度きりです。
幼若なB細胞はまず「D 群 と JH 群」から遺伝子を選び、次に「VH 群」から遺伝子を選びます。
最初に作る重鎖(H鎖)の「CH 群」からは「μ 鎖(ミュー)」の遺伝子しか選べません。
「選ばれなかった遺伝子」のうち、選ばれた遺伝子と選ばれた遺伝子の間にある部分は切り出されます(・・・細部不明な部分もあります)。
最終的には、各群からひとつずつ選ばれた遺伝子が残ります。
ここでは選ばれた遺伝子を「VH1-D1-JH1-CH1」と表現しています。
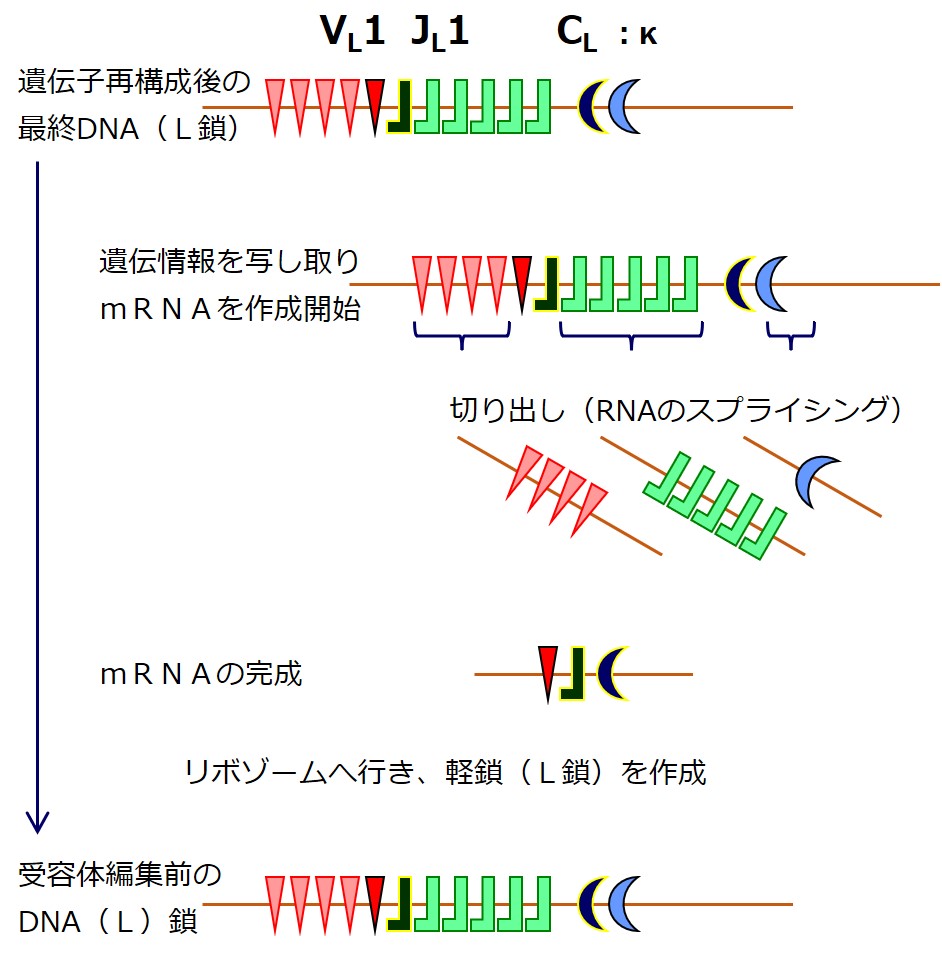
重鎖(H鎖)の遺伝子DNAの再構成が完了したならば、その塩基配列がmRNA(メッセンジャーRNA)に写し取られます。
最初に作られたmRNAから、直接タンパク合成に利用されない余分な配列が切り出され、シンプルになってからリボゾームでのタンパク合成の設計図として使われます。
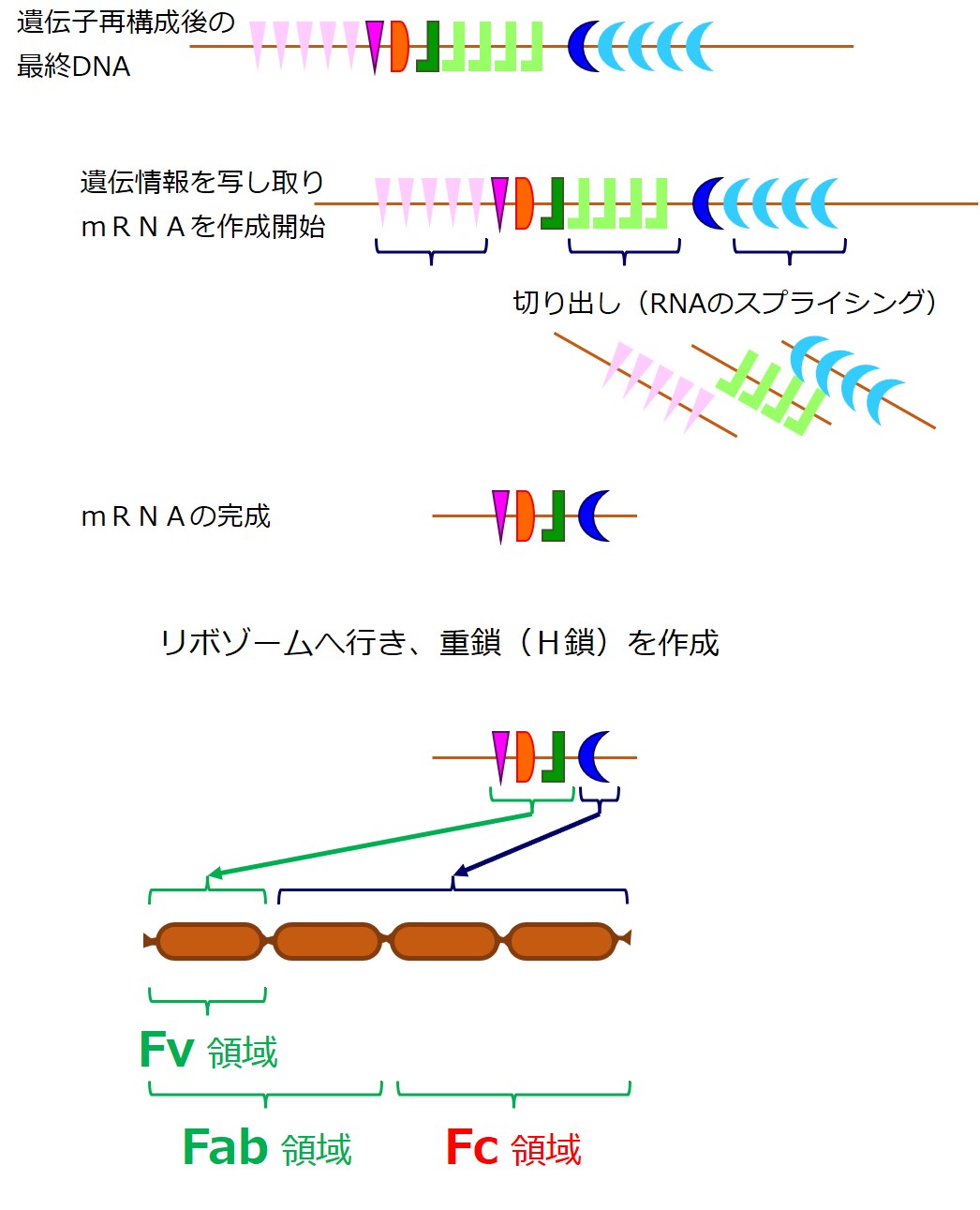
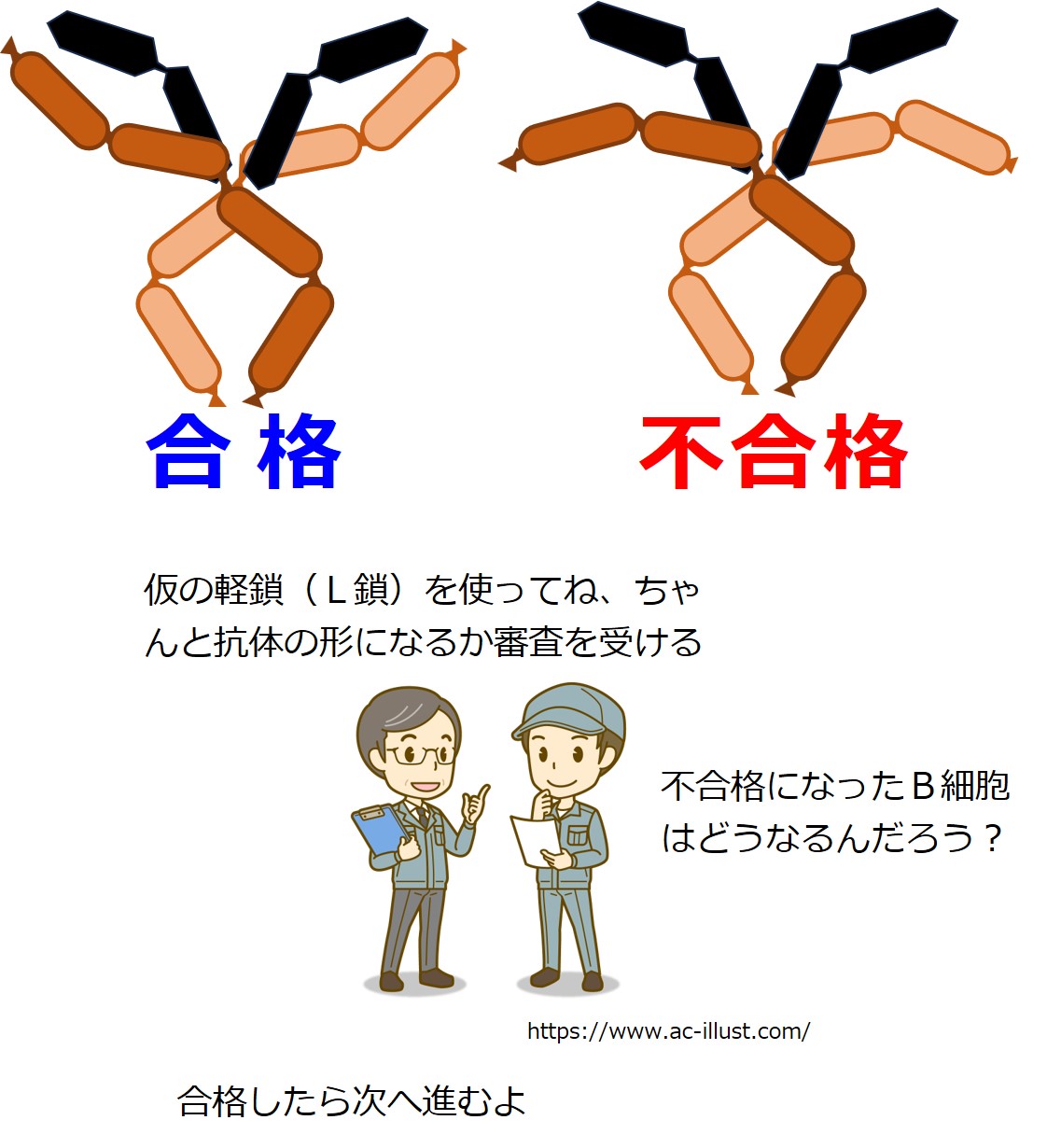
次は、ちゃんとした重鎖(H鎖)ができたかどうかの審査を受けます。
審査は抗体を使っておこなわれます。
重鎖(H鎖)だけでは、抗体ではありません。
しかしまだ、軽鎖(L鎖)は作られていません。
そこであらかじめ用意可能な「偽軽鎖(仮の軽鎖)」を使うそうです。
重鎖(H鎖)2本と偽軽鎖(偽L鎖)2本とを合体させたものが、抗体の形になるかどうかをチェックするようです。
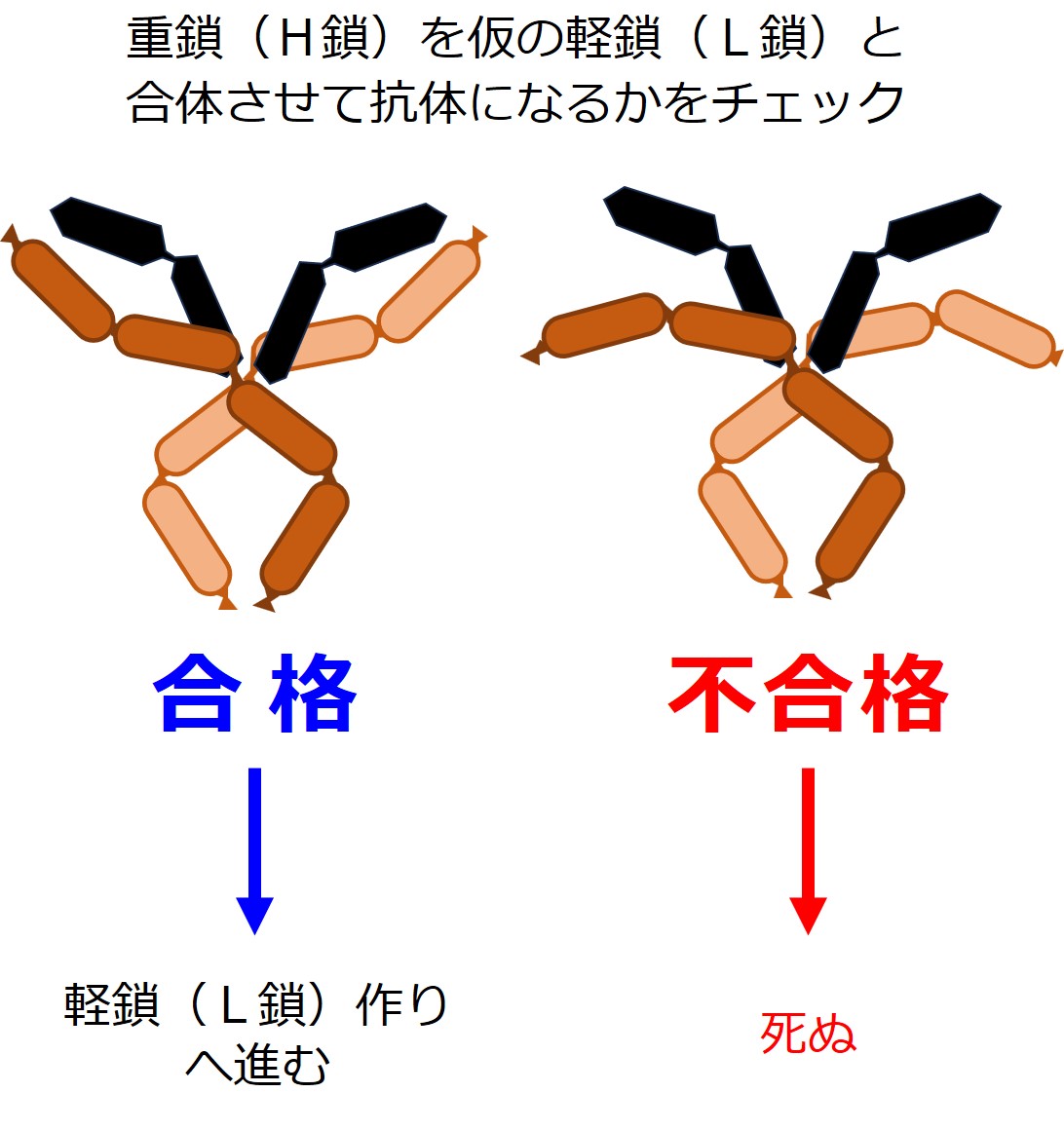
審査方法の詳細は不明ですが、抗体の形にならなかった場合、この重鎖(H鎖)を産生するB細胞は殺されます。
重鎖(H鎖)を作り直すチャンスは与えられないようです。
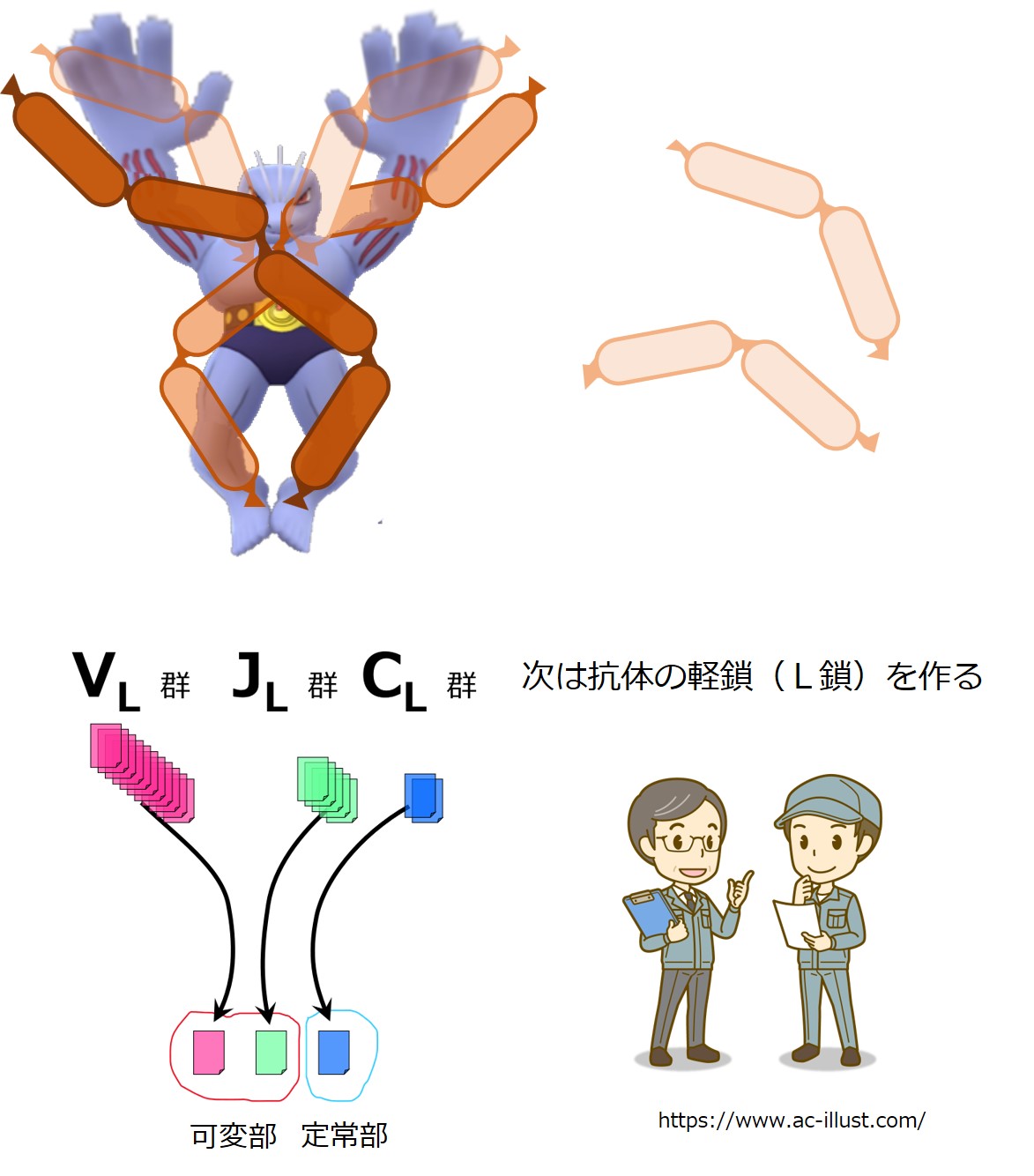
重鎖(H鎖)の合格したB細胞は、軽鎖(L鎖)作りを始めます。
軽鎖(L鎖)の遺伝子は、{V、D、J、C}の各遺伝子群に分かれています。
軽鎖(L鎖)にはD群が無いので、軽鎖(L鎖)の遺伝子群は{VL-JL-CL}と表現されることがあります。
CL 群の遺伝子はカッパ(κ)とラムダ(λ)の2種類あります。一定の比率でどちらかが選ばれるようです。
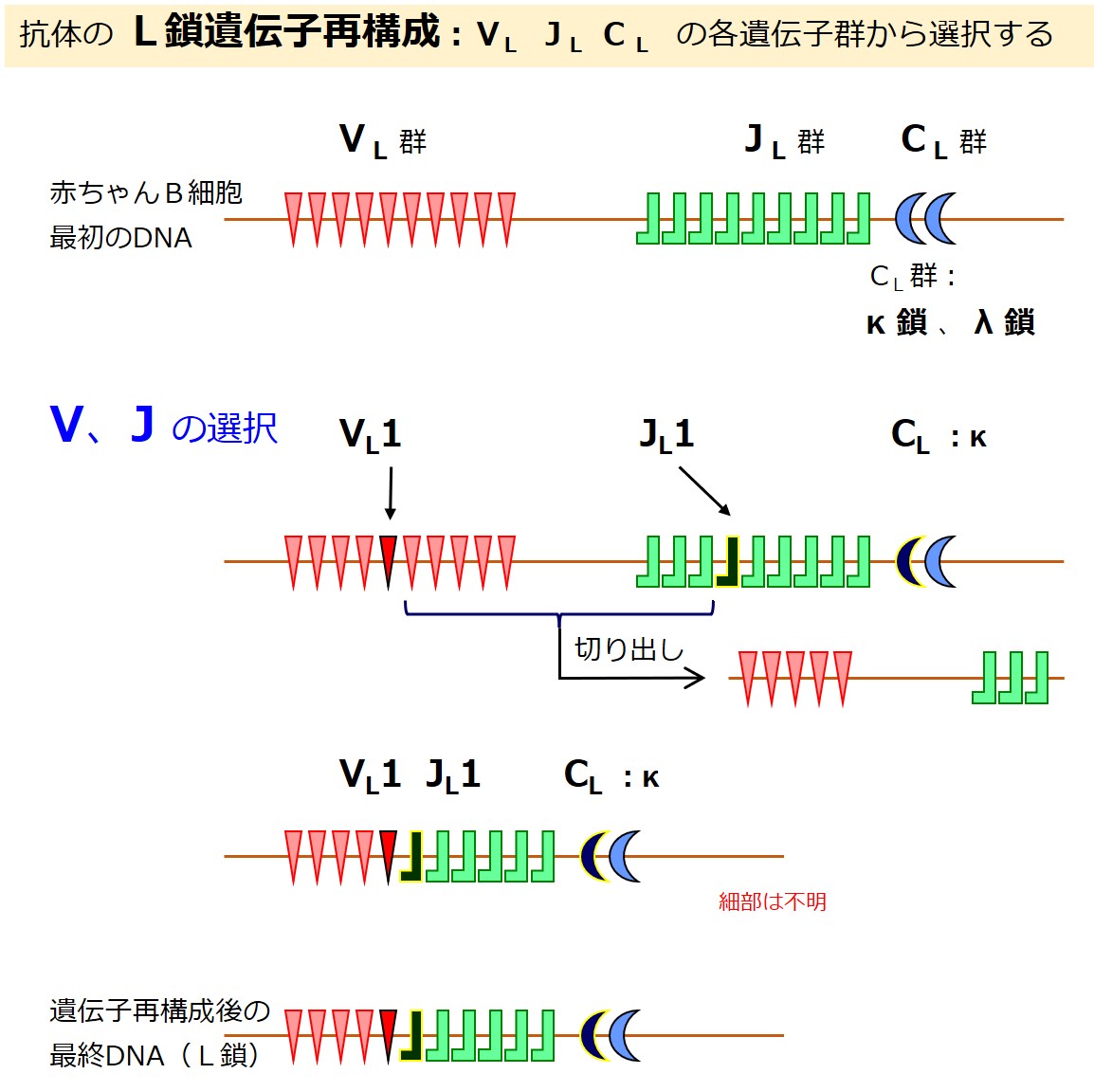
遺伝子再構成により軽鎖(L鎖)の遺伝子が決まると、軽鎖(L鎖)の産生を開始します。
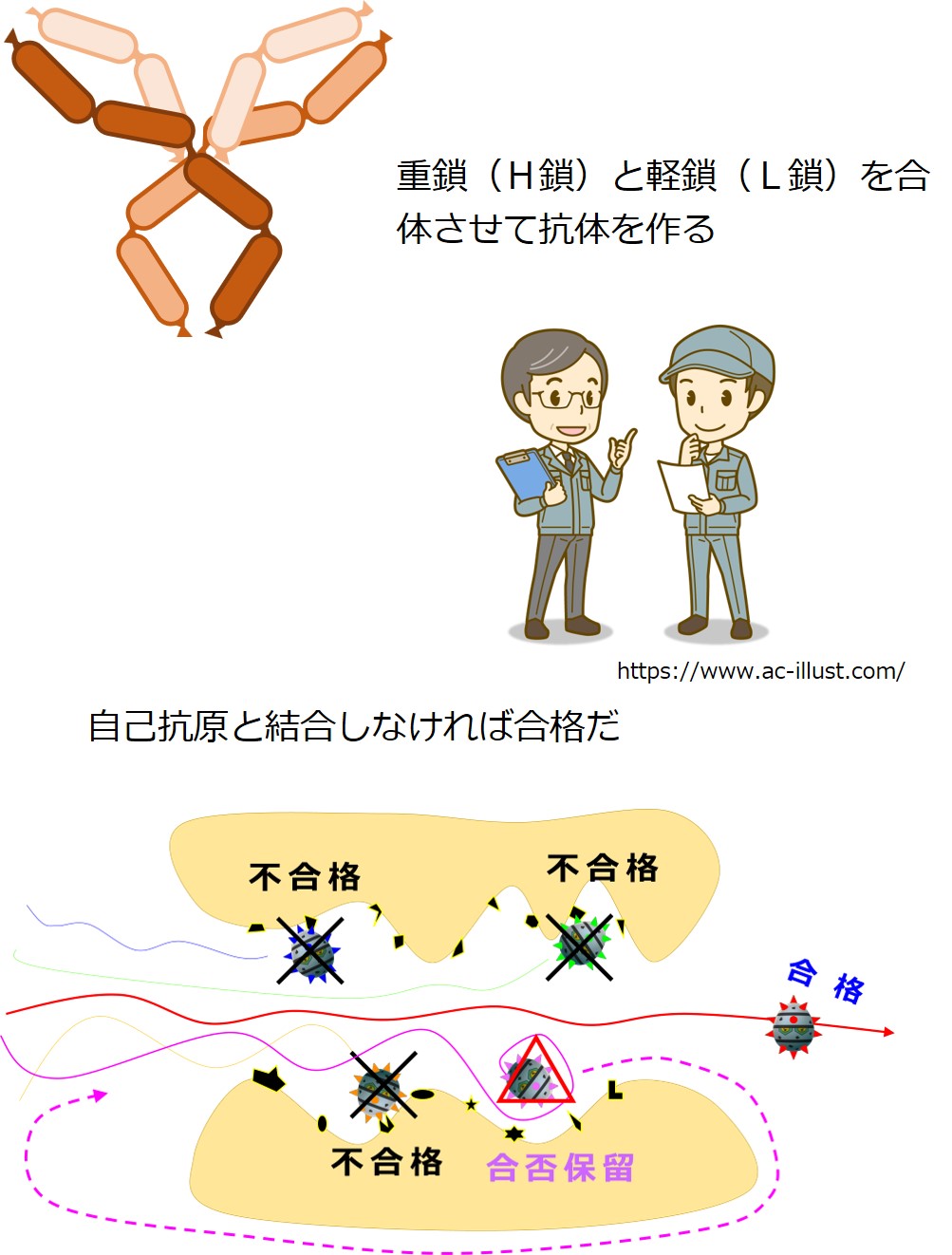
重鎖(H鎖)と軽鎖(L鎖)を合体させて抗体を産生します。
このころの抗体はB細胞の表面に出ています。B細胞受容体(B細胞膜抗原レセプター)と呼ばれる段階です。
自分のオリジナル抗体を産生できるようになった幼若なB細胞は、適切な抗体を産生しているかどうかチェックを受けます。
骨髄の中には全身のあらゆる自己抗原を提示している細胞領域があり、幼若なB細胞がそこを通過します。
いかなる自己抗原とも結合しないで通過できれば「合格」です。
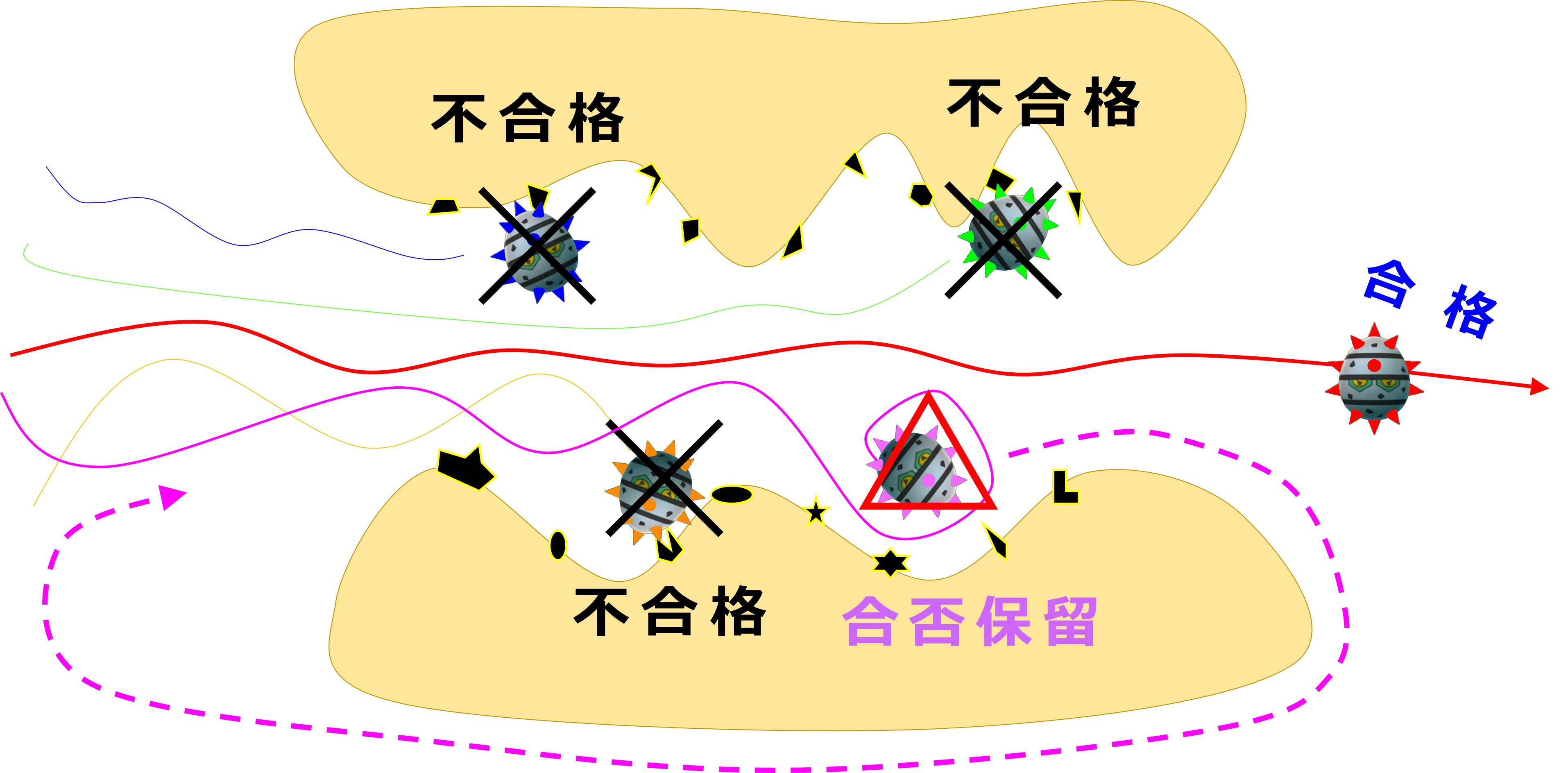
もし自己抗原とピタッと結合した場合は不合格となり、殺されます。
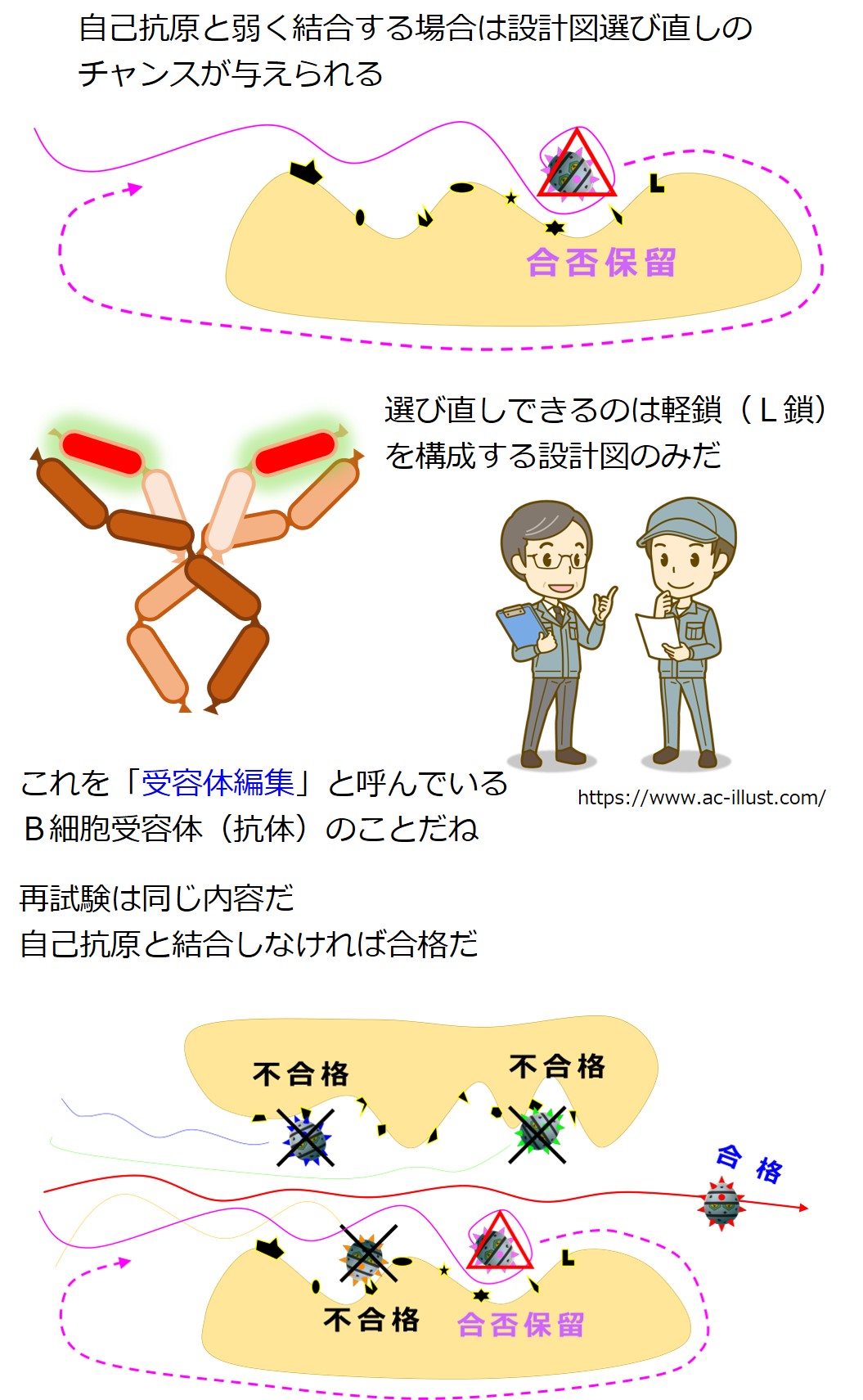
もし自己抗原と結合するかしないか、態度のあいまいな抗体を産生しているB細胞は、抗体を作り直すチャンスが与えられます。
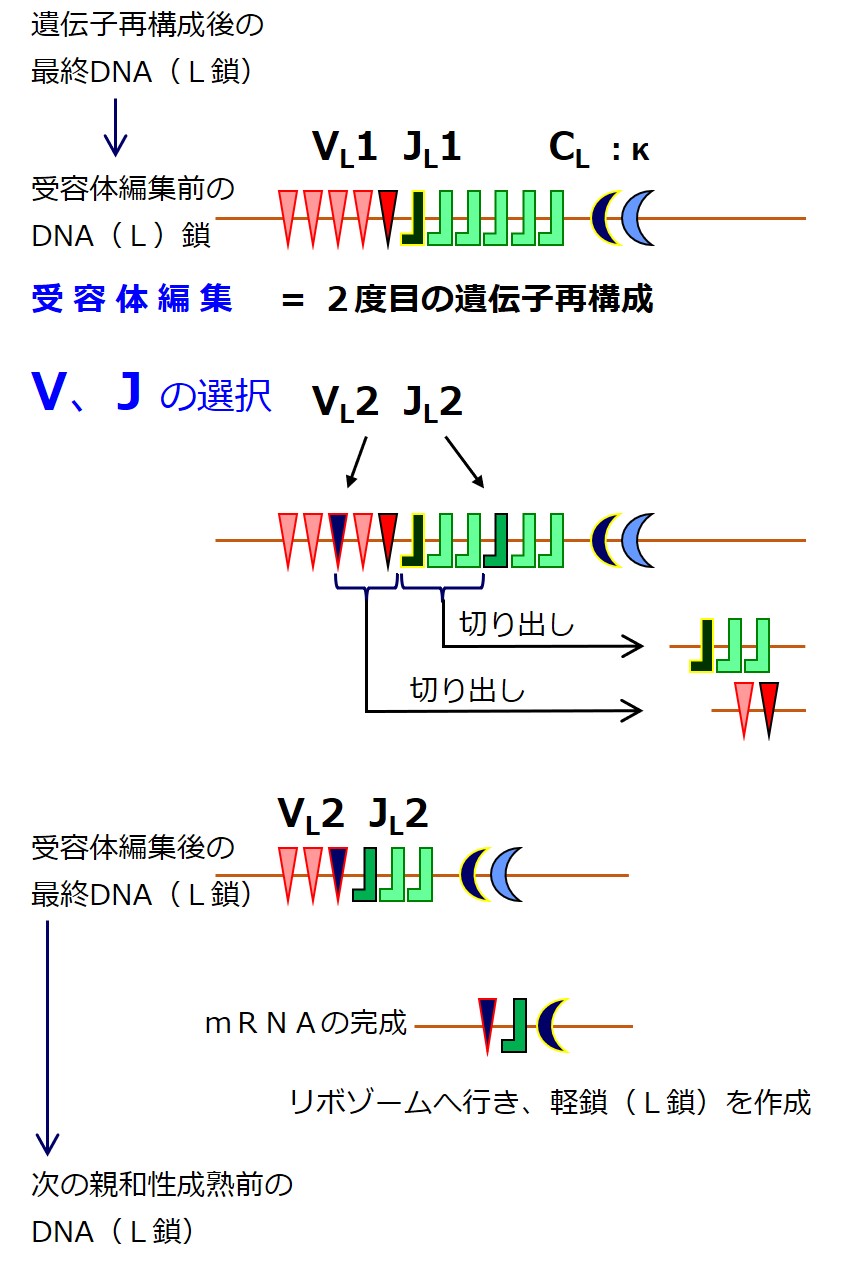
これを受容体編集と呼びます。
この段階での抗体の作り直しは「遺伝子の選び直し」であり、軽鎖(L鎖)のみに許されています。
(注意:リンパ節での親和性成熟における抗体の作り直しは、遺伝子の突然変異であり、軽鎖と重鎖の両方でおこります。骨髄の受容体編集における抗体の作り直しは、遺伝子の選び直しであり、軽鎖のみでおこります。)
遺伝子群(VL 群 と JL 群)から遺伝子を選び直すのですが、最初の遺伝子再構成のときに、選ばれた2つの遺伝子の間にある「選ばれなかった遺伝子」は切り出されて失われています。
「選ばれなかった遺伝子」であっても、選ばれた2つの遺伝子に挟まれていなければ残っています。
最初の遺伝子再構成のときよりも選択肢はぐっと少なくなっています。そのあたりを図でしっかりと理解しましょう。
受容体編集後は、自己抗原との結合性についてふたたびチェックを受けます。
合格だと骨髄を出て旅立ちます、不合格だと殺され、合否保留だとまた受容体編集(骨髄での3度目の遺伝子再構成)に進みます。
受容体編集を繰り返すごとに、遺伝子群の遺伝子の種類は減っていきます。
そのために合格が難しくなるかどうかはわかりません。
次は、骨髄を卒業して全身を巡回しているB細胞を追いましょう。
この段階のB細胞は、IgM 抗体をB細胞受容体として細胞膜上に出しています。
さて、けっこう手ごわい病原体が感染しました。
病原体を食べた樹状細胞は少し活性化します。
活性化と同時に樹状細胞の寿命は残り数日と短くなります。
死のタイマーがセットされると言うそうです。
死のタイマーは、強力な獲得免疫の暴走を防ぐ仕組みのひとつのようです。
樹状細胞は最寄りのリンパ節に移動し、幼若なT細胞に対して食べた病原体の抗原を提示します。
その病原体の抗原を標的として担当する幼若なヘルパーT細胞に対して抗原提示が成功すると、ヘルパーT細胞は活性化します。

活性化したヘルパーT細胞は増殖します。
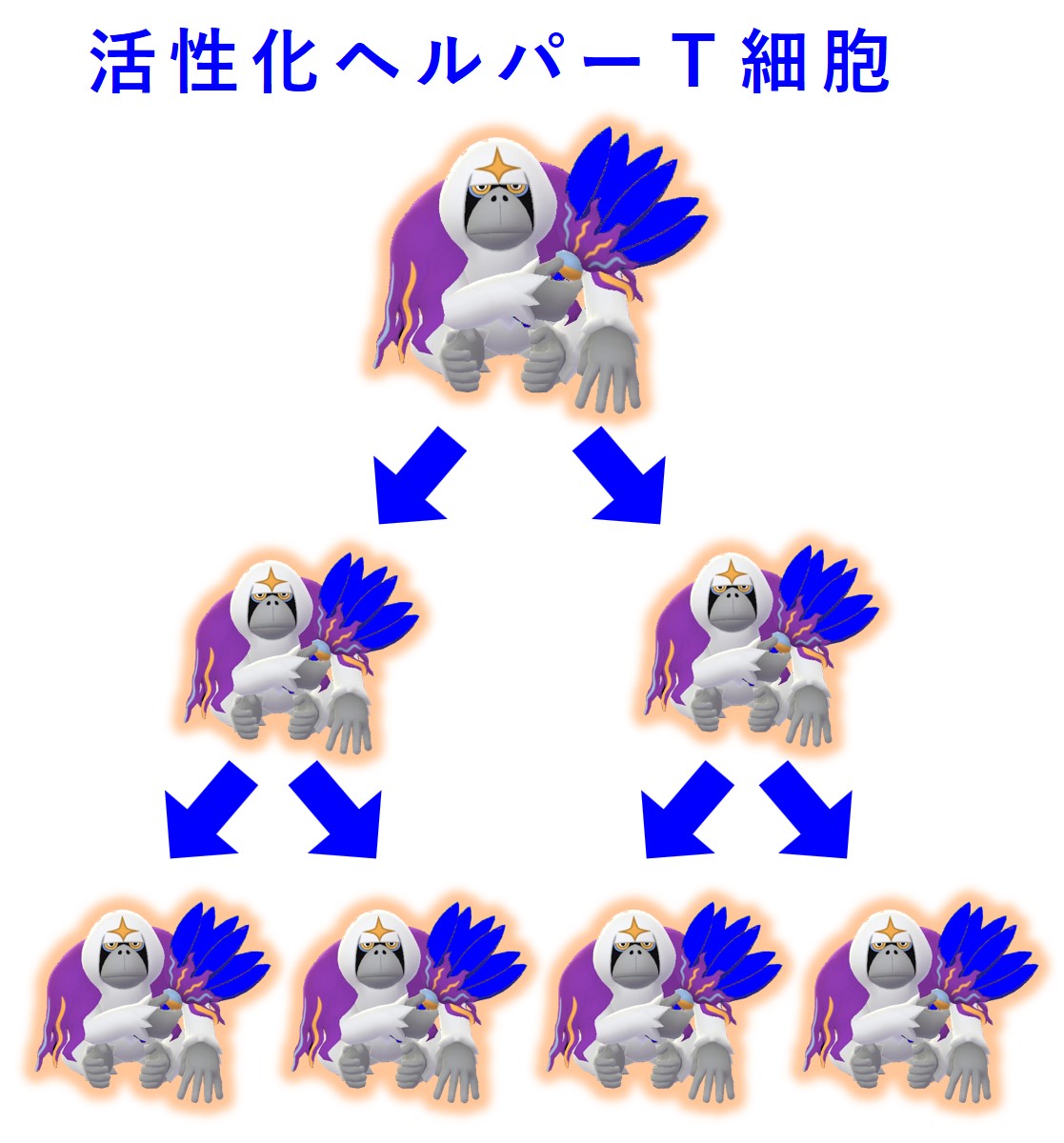
増殖した活性化ヘルパーT細胞の一部はリンパ節に残り、同じ病原体の抗原を標的として担当する幼若なB細胞を探します。
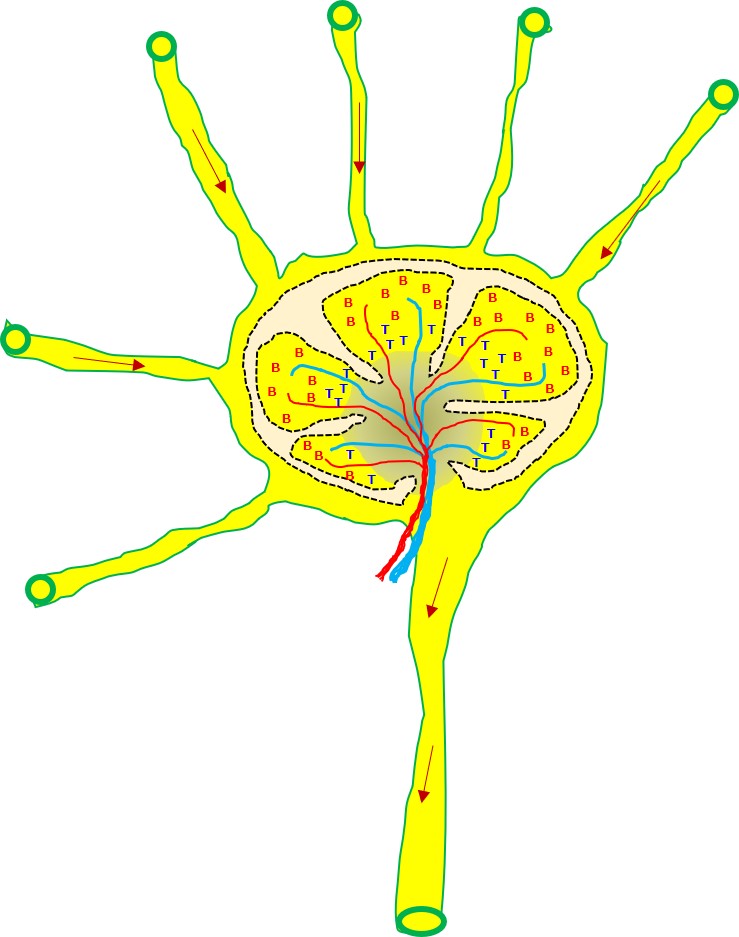
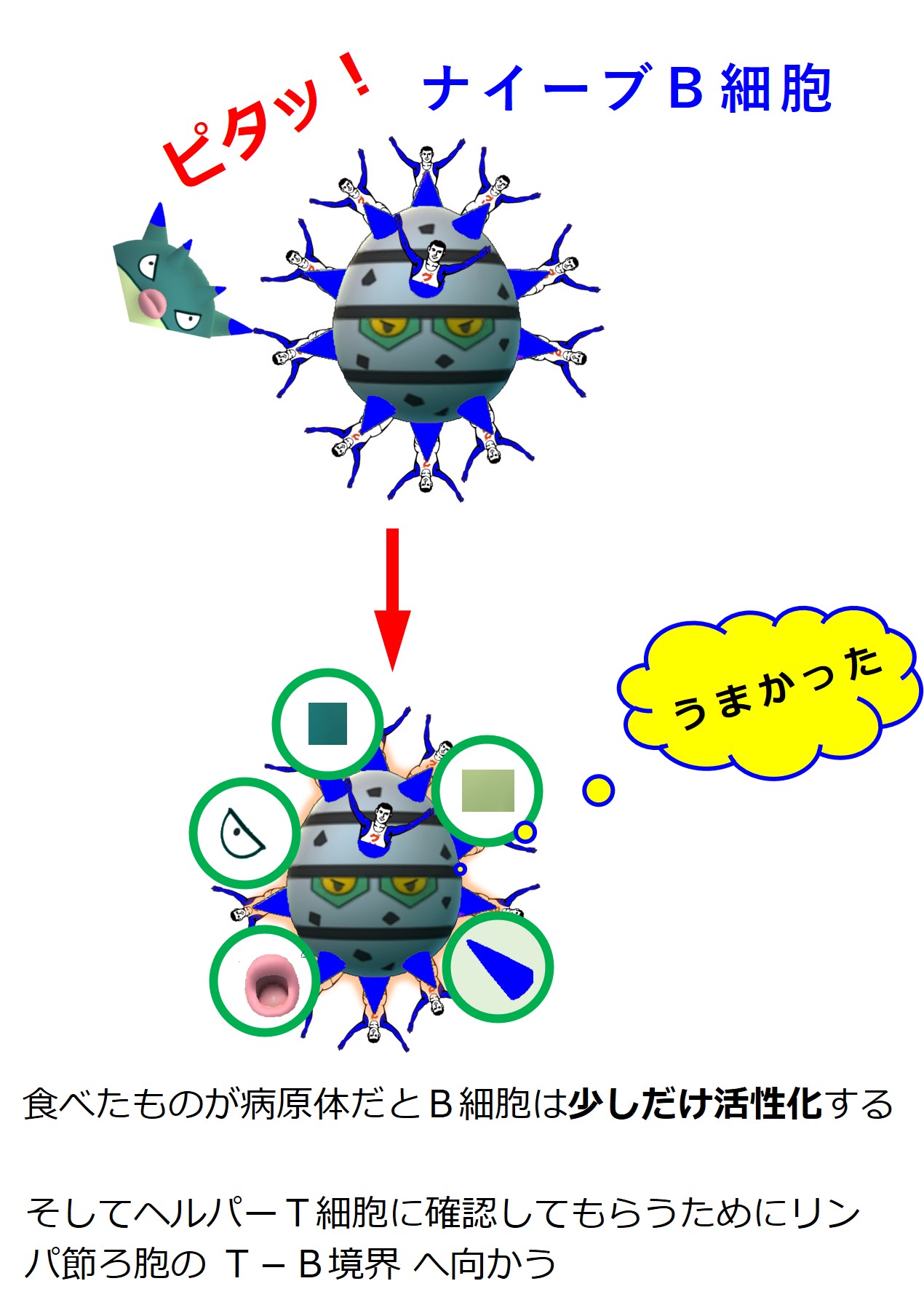
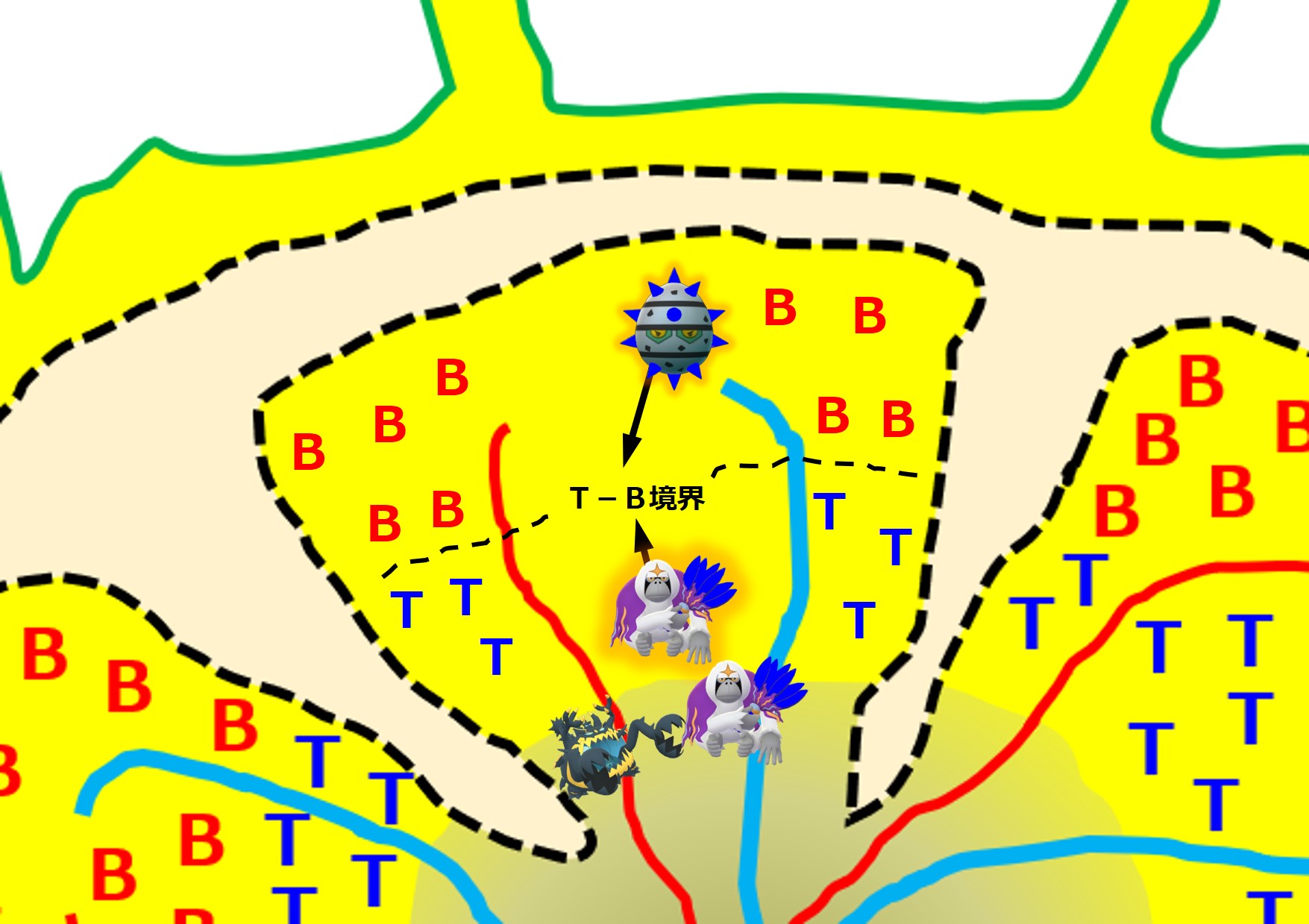
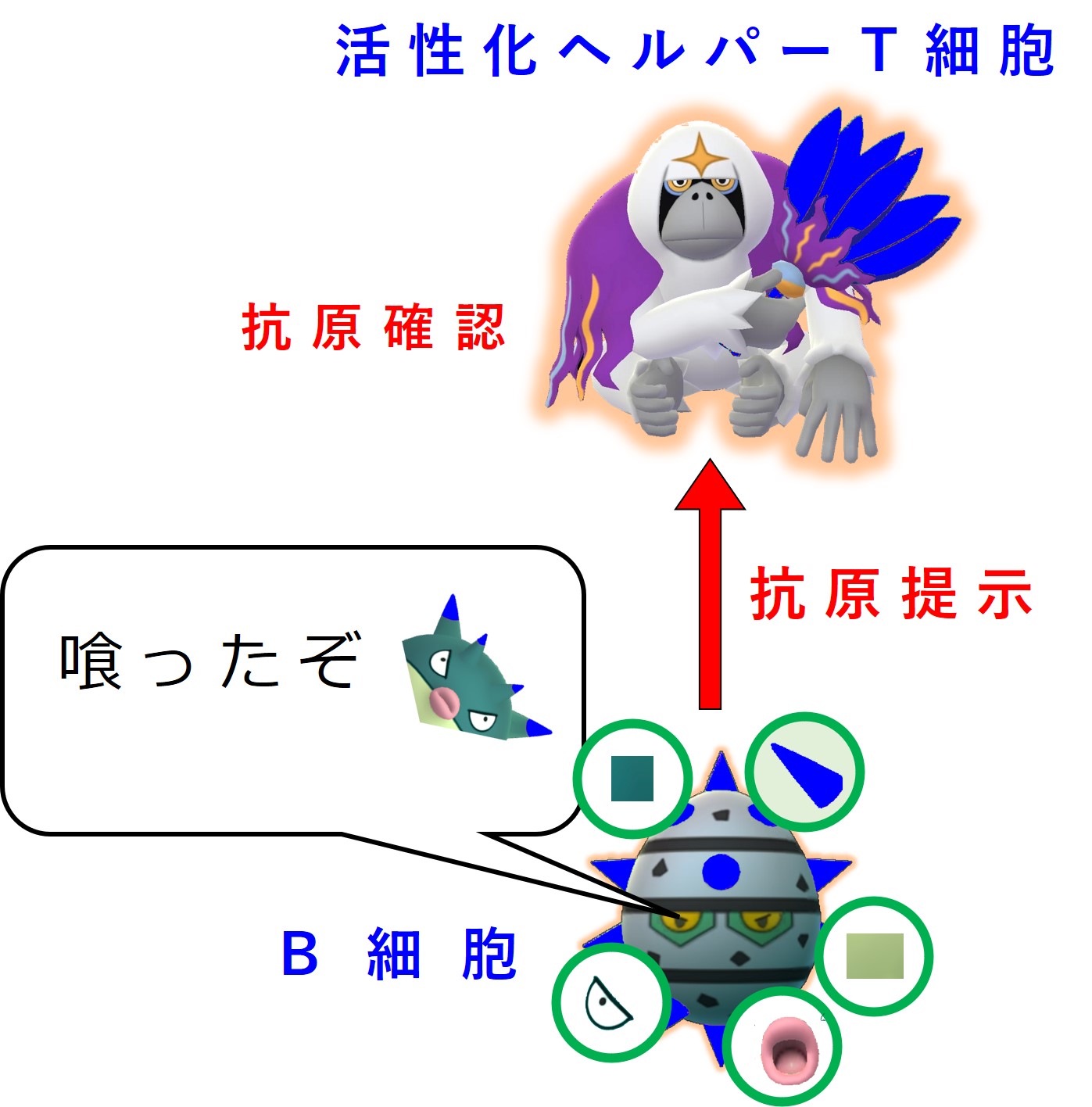
ヘルパーT細胞は、同じ病原体にあるエピトープに結合する抗体(B細胞受容体)を持つB細胞を見つけると、このB細胞を活性化します。
図ではグリコのおじさんを省略していますが、トゲトゲがB細胞受容体(抗体)を意味しています。
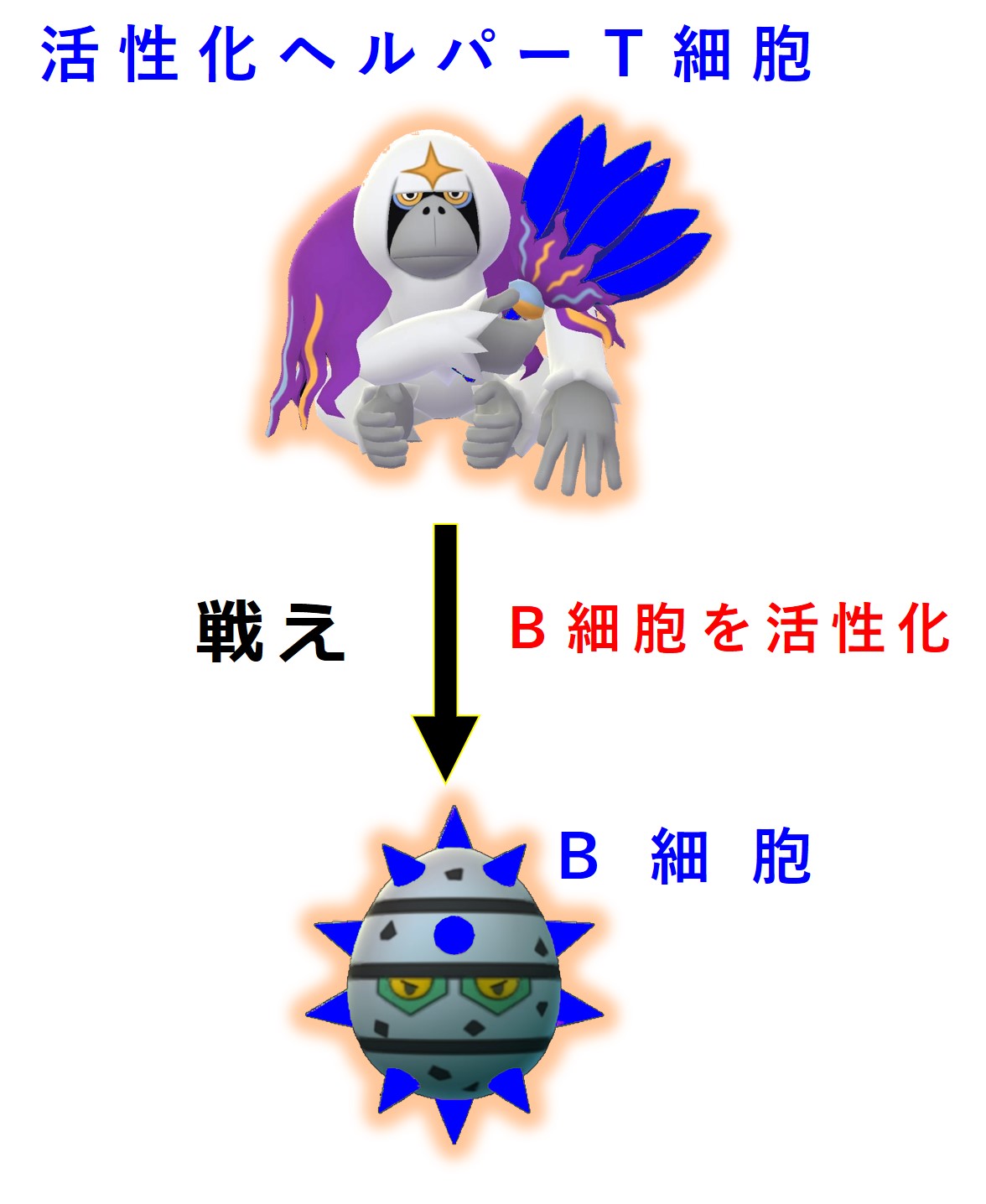
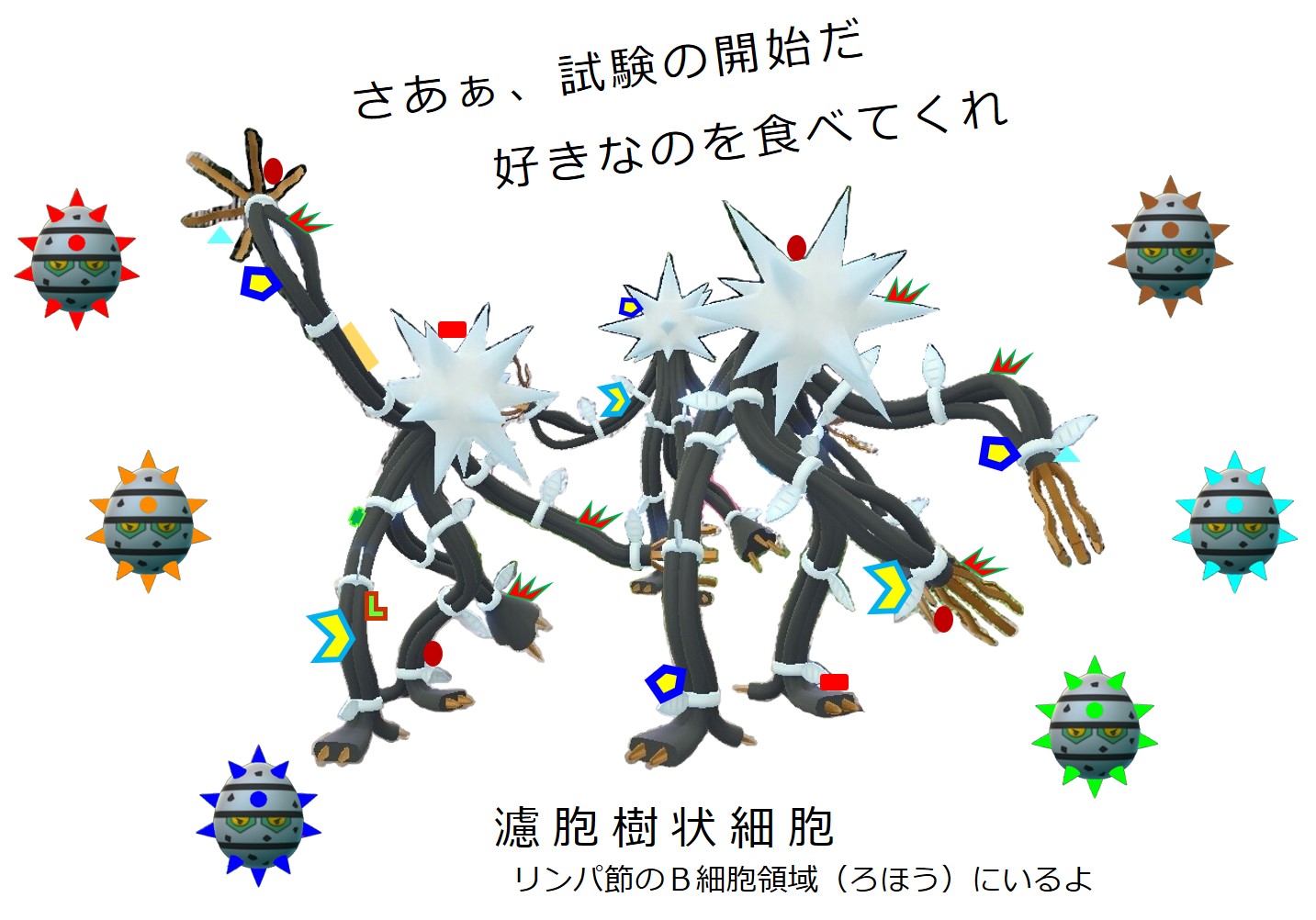
ヘルパーT細胞によってお墨付きをもらって活性化したB細胞に起こることが「3-1-2-4.親和性成熟とクラススイッチ」で説明している 第1選別 です。
リンパ節における最初の試練ですが、この段階では殺されるB細胞はいません。
自己抗原に結合するB細胞はいないからです。
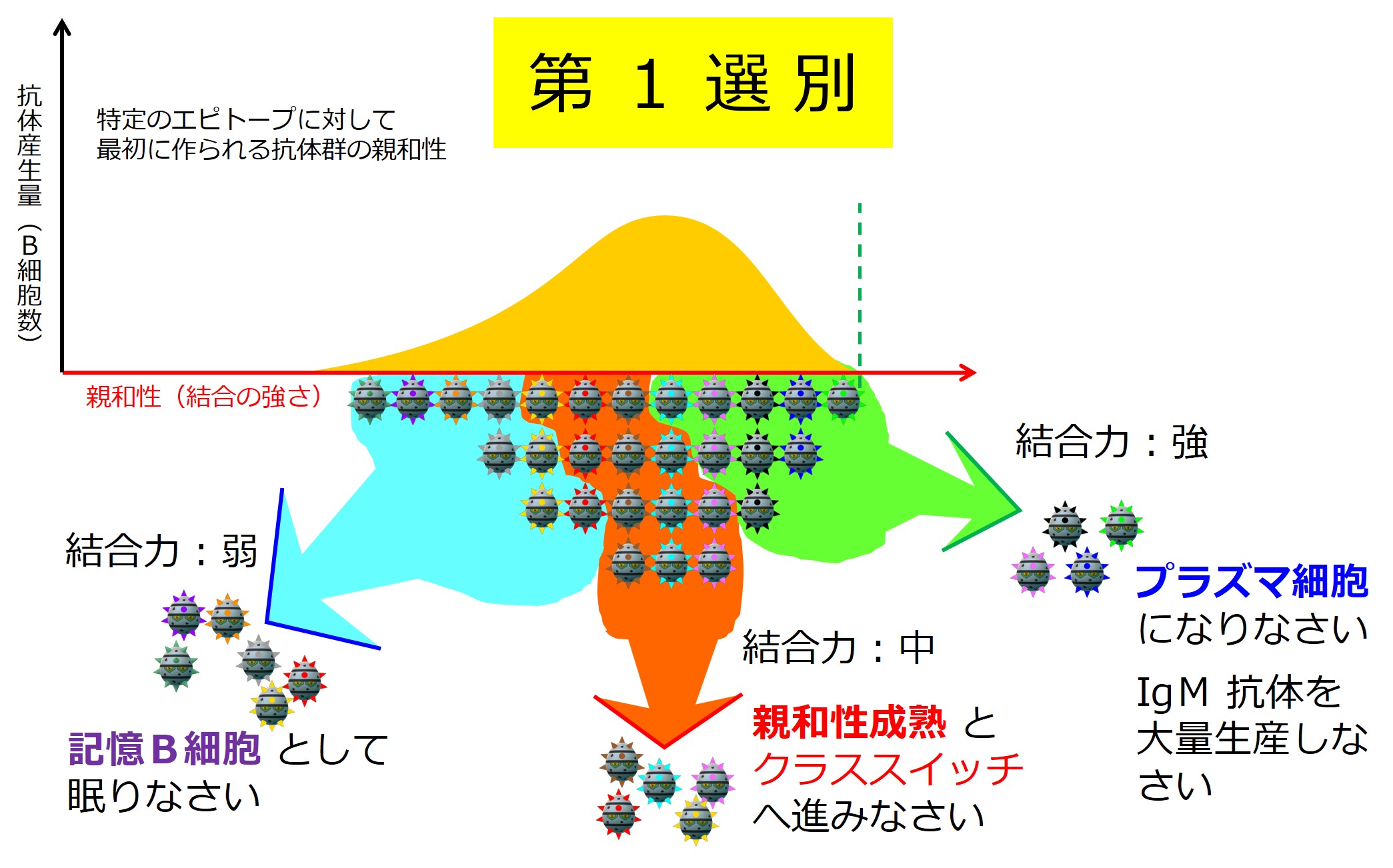
| B細胞の第1選別(リンパ節) | |
| 非自己抗原との結合 | 行き先 |
| +++ | プラズマ細胞 |
| ++ | 親和性成熟 |
| + | メモリーB細胞 |
| ー | ナイーブのまま |
しかし、親和性成熟に進んだB細胞には体細胞超突然変異がおこり、その過程では自己抗原に反応するB細胞も生じるので、次の審査では殺されるB細胞が出ます。
親和性成熟で重鎖(H鎖)と軽鎖(L鎖)の可変部に生じる体細胞超突然変異は、遺伝子の再構成ではありません。DNAの塩基(AGTC)が突然変異で入れ替わり、そのためにアミノ酸が入れ替わり、タンパクが変化します。
親和性成熟と同時期に(詳細不明)、クラススイッチもおこります。クラススイッチは重鎖(H鎖)の遺伝子再構成です。

親和性成熟を終えたB細胞は、ちゃんとした抗体を産生できるようになっているか次の審査を受けます。
突然変異は何が起こるかわかりません。自己抗原に結合する抗体や、何の役にも立たない抗体を産生するように変化した可能性もあり、次の審査は厳しいものになります。
「3-1-2-4.親和性成熟とクラススイッチ」で説明している 第2選別と第3選別 です。
| B細胞の第2選別(リンパ節) | ||
| 割合 | ろ胞樹状細胞の 展示抗原との結合 |
行き先 |
| 1/3 | +++ ~ ++ | 第3選別へ |
| 2/3 | + ~ ー | 死滅 |
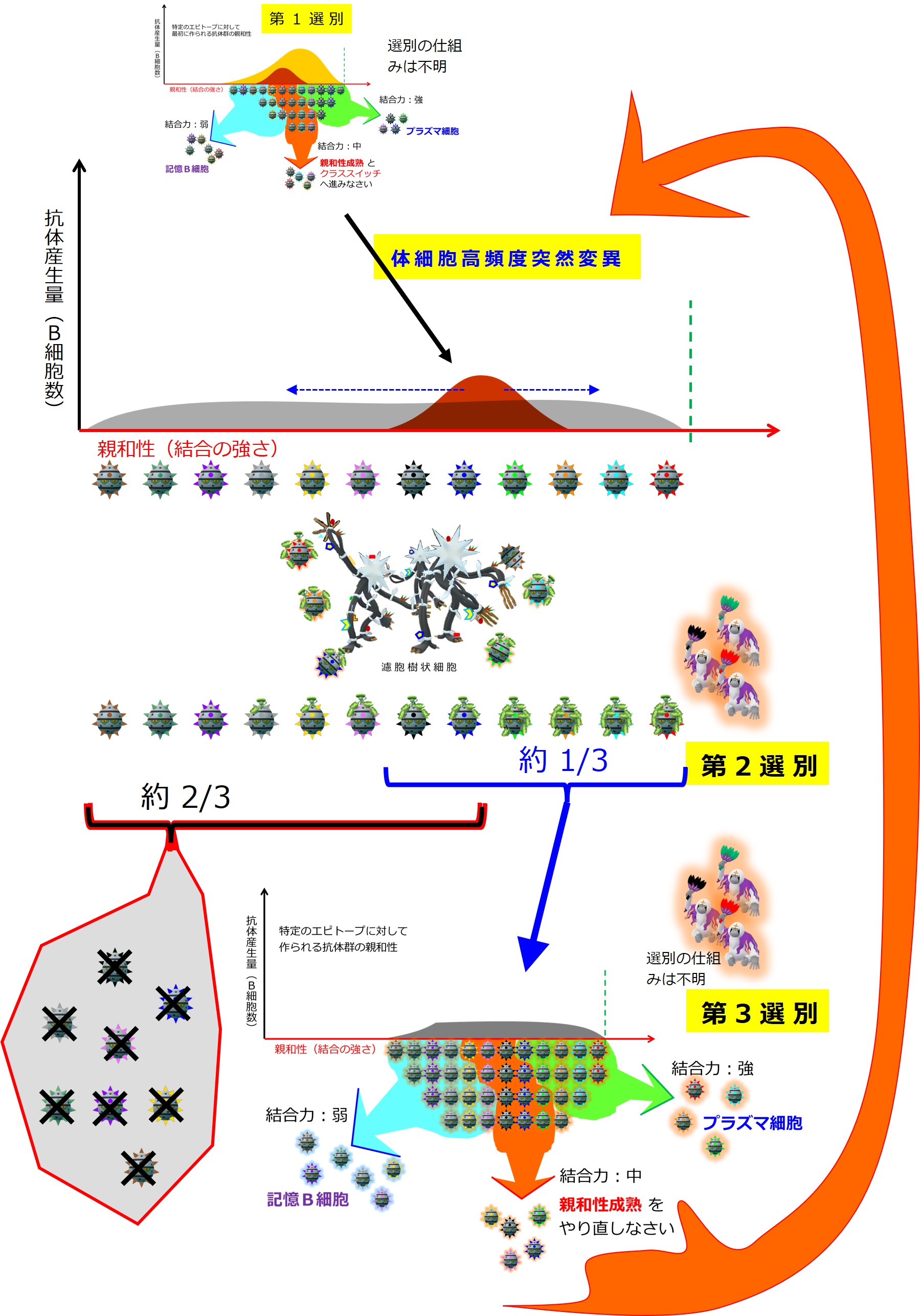
| B細胞の第3選別(リンパ節) | ||
| 割合 | 非自己抗原との結合 | 行き先 |
| 1/3 | +++ | プラズマ細胞 |
| 1/3 | ++ | 親和性成熟やり直し |
| 1/3 | + | メモリーB細胞 |
親和性成熟のやり直しとなったB細胞は同じことを繰り返します。
感染症がピークを越えて、病原体が減ってくると親和性成熟もできなくなります。
以上が、「B細胞が1人前の抗体産生細胞になるまでに受ける試練」の内容です。
なぜこのような厳しい選抜がおこなわれるのか。
大事なことなので繰り返しますが、「免疫細胞とは凶悪な細胞たちです。ゲテモノ食い、殺人鬼、爆破マニアたちです。免疫細胞の様々な生成物には猛毒など強力な殺人兵器が含まれています。抗体(特にIgG)は、敵の目印をつけ、免疫細胞や生成物質による多彩で強力な攻撃を誘導します。ヒトの身体を構成する正常な細胞(自己抗原)を間違って敵と認識するような抗体を作ると大変なことになります。そこで、抗体には厳しい選抜試験が課せられます。」

遺伝子の変化がわかるようになったところで「3-1-2-4.親和性成熟とクラススイッチ」を読み直すと良いでしょう。
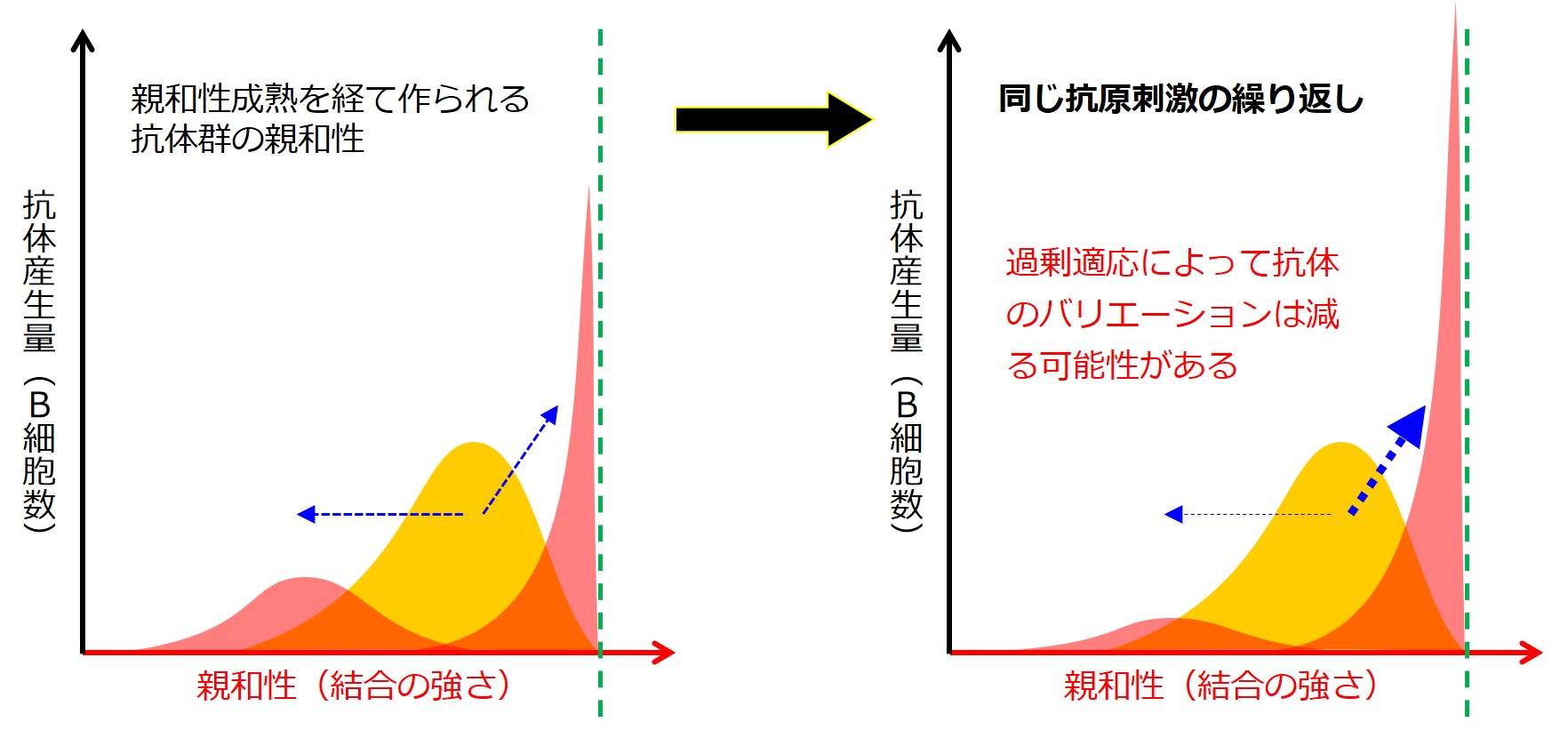
親和性成熟は病原体の変異にも前もって準備していく巧みな仕組みです。
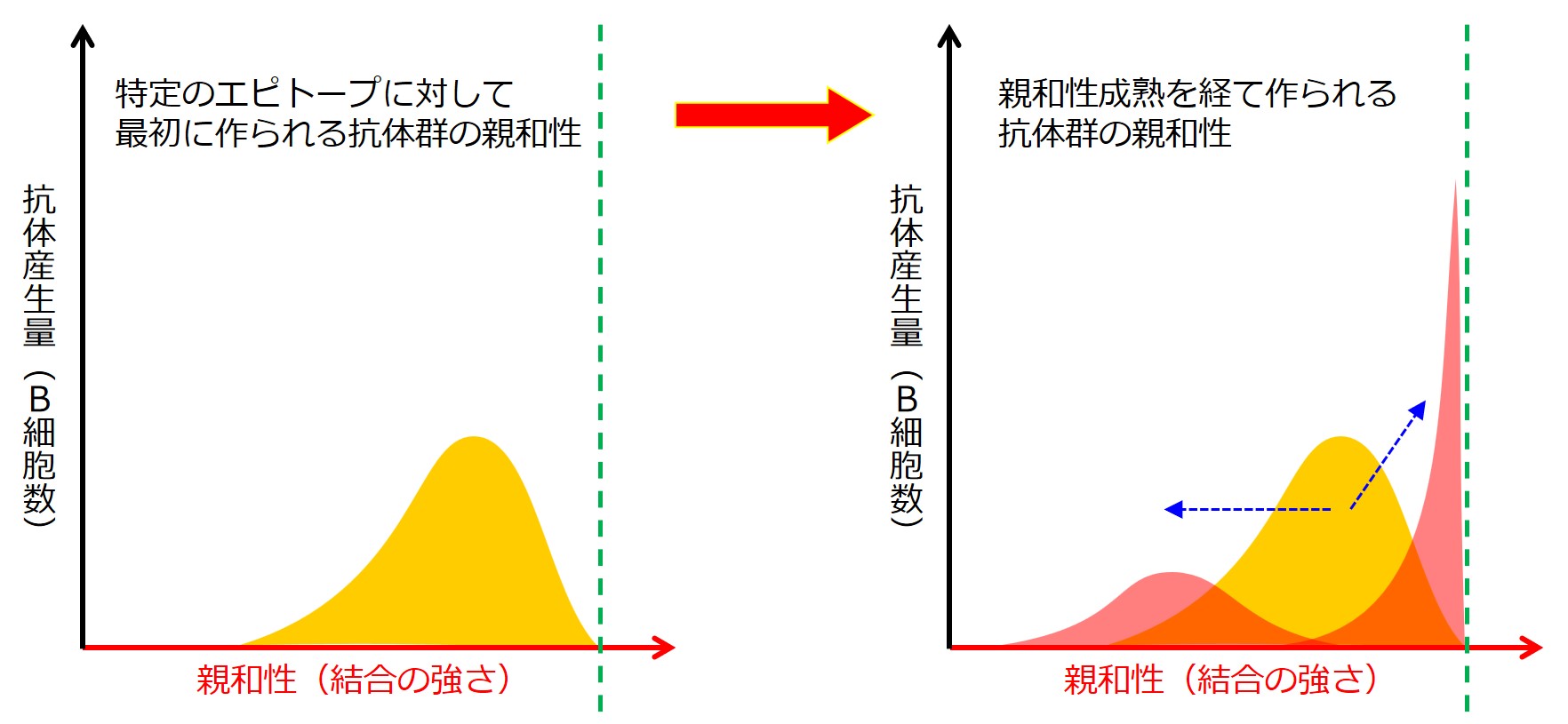
しかし同じ抗原による強烈な刺激を繰り返し受け続けると、過剰適応が生じ、親和性成熟で得られる幅広く柔軟な抗体の準備も損なわれる可能性があります。
同じ抗原によるmRNAワクチン接種(初期型)の繰り返し接種は、親和性成熟を損なった可能性が考えられます。